Basic principles nof metabolism: catabolism, anabolism. Common pathways of proteins, ncarbohydrates and lipids transformation.
Studying of Krebs cycle functioning
INVESTIGATION OF BIOLOGICAL nOXIDATION, OXIDATIVE nPHOSPHORYLATION AND ATP SYNTHESIS. INHIBITORS AND UNCOUPLERS OF OXIDATIVE PHOSPHORYLATION.

Metabolism is the set of life-sustaining chemical transformations within the cells of nliving organisms. These enzyme-catalyzed nreactions allow organisms to grow and reproduce, maintain their structures, and nrespond to their environments. The word metabolism can also refer to all nchemical reactions that occur in living organisms, including digestion and the ntransport of substances into and between different cells, in which case the set nof reactions within the cells is called intermediary metabolism or intermediate nmetabolism.
The term metabolism is derived from the Greek Μεταβολισμός – n”Metabolismos” for “change”, or “overthrow”. The history of the scientific study of nmetabolism spans several centuries and has moved from examining whole animals nin early studies, to examining individual metabolic reactions in moderbiochemistry. The first controlled experiments in human metabolism were published by Santorio nSantorioin
In these early studies, the mechanisms of these nmetabolic processes had not been identified and a vital force was thought to animate living tissue. In the 19th century, when studying the fermentation of nsugar to alcohol by yeast, Louis Pasteur concluded that fermentation was ncatalyzed by substances within the yeast cells he called “ferments”. nHe wrote that “alcoholic fermentation is an act correlated with the life nand organization of the yeast cells, not with the death or putrefaction of the ncells.”] This discovery, along with the npublication by Friedrich nWöhler in 1828 of nthe chemical synthesis of urea, notable for being the first organic ncompound prepared from wholly inorganic precursors, proved that the organic ncompounds and chemical reactions found in cells were no different in principle nthan any other part of chemistry.
Metabolism is a term that is used to describe all nchemical reactions involved in maintaining the living state of the cells and nthe organism. Metabolism can be conveniently divided into two categories:
· nCatabolism n- the breakdown of molecules to obtain energy
· nAnabolism – the synthesis of all compounds nneeded by the cells
Anabolism is the set of constructive metabolic processes where the nenergy released by catabolism is used to synthesize complex molecules. Igeneral, the complex molecules that make up cellular structures are constructed nstep-by-step from small and simple precursors. Anabolism involves three basic nstages. Firstly, the production of precursors such as amino acids, monosaccharides,isoprenoids and nucleotides, nsecondly, their activation into reactive forms using energy from ATP, and nthirdly, the assembly of these precursors into complex molecules such as proteins, polysaccharides, lipids and nucleic acids.
Metabolism refers to the highly integrated network of chemical nreactions by which living cells grow and sustain themselves. This network is ncomposed of two major types of pathways: anabolism and catabolism. Anabolism nuses energy stored in the form of adenosine triphosphate (ATP) to build larger nmolecules from smaller molecules. Catabolic reactions degrade larger molecules nin order to produce ATP and raw materials for anabolic reactions.
Together, these two general metabolic networks have nthree major functions:
(1) to extract nenergy from nutrients or solar energy;
(2) to synthesize the building blocks that make up the nlarge molecules of life: proteins, fats, carbohydrates, nucleic acids, and ncombinations of these substances;
(3) to synthesize and degrade molecules required for nspecial functions in the cell.
The series of products created by the sequential nenzymatic steps of anabolismor catabolism are called metabolic intermediates, nor metabolites. Each steprepresents a small change in the molecule, usually the nremoval, transfer, oraddition of a specific atom, molecule, or group of atoms nthat serves as a functional group, such as the amino groups (-NH2) of proteins.
Most such metabolic pathways are linear, that is, they nbegin with a specificsubstrate and end with a specific product. However, some npathways, such as the Krebs cycle, are cyclic. Often, metabolic pathways also nhave branches thatfeed into or out of them. The specific sequences of nintermediates in the pathways of cell metabolism are called intermediary nmetabolism.
Among the many hundreds of chemical reactions there are nonly a few that are central to the activity of the cell, and these pathways are nidentical in mostforms of life.
Catabolic reactions are used to capture and save energy from nutrients, nas well as to degrade larger molecules into smaller, molecular raw materials nfor reuse by the cell. The energy is stored in the form of energy-rich ATP, nwhichpowers the reactions of anabolism. The useful energy of ATP is stored itheform of a high-energy bond between the second and third phosphate groups of ATP. nThe cell makes ATP by adding a phosphate group to the molecule nadenosinediphosphate (ADP). Therefore, ATP is the major chemical link betweethe energy-yielding reactions of catabolism, and the energy-requiring reactions nof anabolism.
In some cases, energy is also conserved as energy-rich hydrogen atoms ithecoenzyme nicotinamide adenine dinucleotide phosphate in the reduced form of nNADPH. The NADPH can then be used as a source of high-energy hydrogen atoms nduring certain biosynthetic reactions of anabolism.
In addition to the obvious difference in the direction of their metabolic ngoals, anabolism and catabolism differ in other significant ways. For example, nthe various degradative pathways of catabolism are convergent. That is, many nhundreds of different proteins, polysaccharides, and lipids are broken dowinto relatively few catabolic end products. The hundreds of anabolic npathways,however, are divergent. That is, the cell uses relatively few nbiosynthetic precursor molecules to synthesize a vast number of different nproteins, polysaccharides, and lipids.
The opposing pathways of anabolism and catabolism may also use different nreaction intermediates or different enzymatic reactions in some of the steps. nForexample, there are 11 enzymatic steps in the breakdown of glucose into npyruvic acid in the liver. But the liver uses only nine of those same steps ithesynthesis of glucose, replacing the other two steps with a different set nofenzyme-catalyzed reactions. This occurs because the pathway to degradatioofglucose releases energy, while the anabolic process of glucose synthesis nrequires energy. The two different reactions of anabolism are required to novercome the energy barrier that would otherwise prevent the synthesis of nglucose.
Another reason for having slightly different pathways is that the ncorresponding anabolic and catabolic routes must be independently regulated. nOtherwise,if the two phases of metabolism shared the exact pathway (only ireverse) aslowdown in the anabolic pathway would slow catabolism, and vice nversa.
Read more: http://www.faqs.org/health/topics/40/Metabolism.html#ixzz2QG2PLkrF
Some reactions can be either catabolic or anabolic, ndepending on the circumstances. Such reactions are called amphibolic reactions. nMany of the reactions interconverting the “simple molecules” fall in this ncategory.
Catabolic and anabolic pathways are interrelated in three ways:
Matter (catabolic pathways furnish the precursor compounds for anabolism. nEnergy (catabolic pathways furnish the energy to “drive” anabolism). Electrons n(catabolic pathways furnish the reducing power for anabolism).
Linear pathways convert one compound through a series of intermediates to nanother compound. An example would be glycolysis, where glucose is converted to npyruvate.
Branched pathways may either be divergent (an intermediate can enter nseveral linear pathways to different end products) or convergent (several nprecursors can give rise to a common intermediate). Biosynthesis of purines and nof some amino acids are examples of divergent pathways. There is usually some nregulation at the branch point. The conversion of various carbohydrates into nthe glycolytic pathway would be an example of convergent pathways.
In a cyclic pathway, intermediates are regenerated, and so some intermediates nact in a catalytic fashion. In this illustration, the cyclic pathway carries nout the net conversion of X to Z. The Tricarboxylic Acid Cycle is an example of na cyclic pathway.
A pool of compounds in equilibrium with each other provides the intermediates nfor converting compounds to a variety of products, depending on what is fed n“into” the pool and what is “withdrawn” from the pool. The phosphogluconate npathway is an example of such a pool of intermediates. The pathway can convert nglucose to CO2, hexoses to pentoses, pentoses to hexoses, pentoses to trioses, netc. depending on what the cell requires in a particular situation. NADPH as a nsource of reducing power for anabolic reactions is also a main product of the nphosphogluconate pathway.
Organisms differ in how many of the molecules in their ncells they can construct for themselves. Autotrophs such as plants can construct the ncomplex organic molecules in cells such as polysaccharides and proteins from nsimple molecules like carbodioxide and water. Heterotrophs, non the other hand, require a source of more complex substances, such as nmonosaccharides and amino acids, to produce these complex molecules. Organisms ncan be further classified by ultimate source of their energy: photoautotrophs nand photoheterotrophs obtain energy from light, whereas chemoautotrophs and nchemoheterotrophs obtain energy from inorganic oxidation reactions.
Metabolism is closely linked to nutrition and the navailability of nutrients. Bioenergetics is a term which describes the nbiochemical or metabolic pathways by which the cell ultimately obtains energy. nEnergy formation is one of the vital components of metabolism.
Carbohydrates in metabolism
Foods supply carbohydrates in three forms: starch, sugar, nand cellulose (fiber). Starches and sugars form major and essential sources of nenergy for humans. Fibers contribute to bulk in diet.
Body tissues depend on glucose for all activities. nCarbohydrates and sugars yield glucose by digestion or metabolism.Most people consume naround half of their diet as carbohydrates.
Proteins in metabolism
Proteins are the main tissue builders in the body. They nare part of every cell in the body. Proteins help in cell structure, functions, nhaemoglobin formation to carry oxygen, enzymes to carry out vital reactions and na myriad of other functions in the body. Proteins are also vital in supplying nnitrogen for DNA and RNA genetic material and energy production.
Fat in metabolism
Fats are concentrated sources of energy. They produce ntwice as much energy as either carbohydrates or protein on a weight basis.
Carbohydrate catabolism is the breakdown of ncarbohydrates into smaller units. Carbohydrates are usually taken into cells nonce they have been digested intomonosaccharides. Once inside, the major route of nbreakdown is glycolysis, nwhere sugars such as glucose and fructose are converted into pyruvate and some ATP is generated.
Pyruvate is aintermediate in several metabolic pathways, but the majority is converted to acetyl-CoA and fed into the citric acid ncycle. Although some more ATP is generated in the citric acid cycle, nthe most important product is NADH, which is made from NAD+ as the acetyl-CoA is oxidized. This noxidation releases carbodioxide as a waste nproduct. In anaerobic conditions, glycolysis produces lactate, nthrough the enzyme lactate dehydrogenase re-oxidizing NADH to NAD+ for re-use nin glycolysis. An alternative route for glucose breakdown is the pentose phosphate pathway, which reduces nthe coenzyme NADPH and produces pentose sugars such asribose, the nsugar component of nucleic acids.
Fats are catabolised by hydrolysis to free fatty acids and glycerol. The nglycerol enters glycolysis and the fatty acids are broken down bybeta noxidation to release nacetyl-CoA, which then is fed into the citric acid cycle. Fatty acids release nmore energy upon oxidation than carbohydrates because carbohydrates contaimore oxygen in their structures.
Amino acids are neither used to synthesize proteins and other biomolecules, or oxidized to urea and carbon dioxide as a source of nenergy. The oxidation pathway nstarts with the removal of the amino group by a transaminase. nThe amino group is fed into the urea cycle, nleaving a deaminated carbon skeleton in the form of a keto acid. nSeveral of these keto acids are intermediates in the citric acid cycle, for nexample the deamination of glutamate forms α-ketoglutarate. The glucogenic amino acids can also be converted into glucose, nthrough gluconeogenesis .
Stages of catabolism
Catabolism can be broken down into 3 main stages.
Stage 1 – Stage of Digestion
The large organic molecules like proteins, lipids and polysaccharides are digested into ntheir smaller components outside cells. This stage acts on starch, cellulose or nproteins that cannot be directly absorbed by the cells and need to be brokeinto their smaller units before they can be used in cell metabolism.
Digestive enzymes include glycoside hydrolases that ndigest polysaccharides into monosaccharides or simple sugars.
The primary enzyme involved in protein digestion is npepsin which catalyzes the nonspecific hydrolysis of peptide bonds at aoptimal pH of 2. In the lumen of the small nintestine, the pancreas secretes zymogens of trypsin, nchymotrypsin, elastase etc. These proteolytic enzymes break the proteins ndown into free amino acids as well as dipeptides and tripeptides. The free namino acids as well as the di and tripeptides are absorbed by the intestinal nmucosa cells which subsequently are released into the blood stream where they nare absorbed by other tissues.
The amino acids and sugars are then pumped into cells by nspecific active transport proteins.
Stage 2 – Release of energy
Once broken down these molecules are taken up by cells nand converted to yet smaller molecules, usually acetyl coenzyme A (acetyl-CoA), nwhich releases some energy.
Stage 3 – The acetyl group on the CoA is oxidised to water and ncarbon dioxide in the citric acid cycle and electron transport chain, releasing nthe energy that is stored by reducing the coenzyme nicotinamide adenine ndinucleotide (NAD+) into NADH.
Carbohydrate breakdown
When complex carbohydrates are broken they form simple sugars or nmonosaccharides. This is taken up by the cells. Once inside these sugars nundergo glycolysis, where sugars such as glucose and fructose are converted ninto pyruvate and some ATP is generated. Pyruvate is an intermediate in several nmetabolic pathways, but the majority is converted to acetyl-CoA and fed into nthe citric acid cycle or the Kreb’s cycle.
Within the citric acid cycle more ATP is generated by nthe monosaccharides. The most important product is NADH, which is made from nNAD+ as the acetyl-CoA is oxidized. This oxidation releases carbon dioxide nas a waste product.
When there is no oxygen, glycolysis produces lactate, nthrough the enzyme lactate dehydrogenase, re-oxidizing NADH to NAD+ for re-use nin glycolysis.
Glucose can also be broken down by pentose phosphate npathway, which reduces the coenzyme NADPH and produces pentose sugars such as nribose, the sugar component of nucleic acids.
Amino acid breakdown
Proteins are broken down into amino acids. Amino acids nare either used to synthesize proteins and other biomolecules, or oxidized to nurea and carbon dioxide as a source of energy.
In the process of oxidation, first the amino group is nremoved by a transaminase. The amino group is fed into the urea cycle, leaving na deaminated carbon skeleton in the form of a keto acid.
These keto acids enter the citric acid cycle. Glutamate, nfor example, forms α-ketoglutarate. Some of the amines may also be nconverted into glucose, through gluconeogenesis.
Lipid breakdown
Fats are catabolised by hydrolysis to free fatty acids nand glycerol. The glycerol enters glycolysis and the fatty acids are brokedown by beta oxidation to release acetyl-CoA. This acetyl co-A reaches the ncitric acid cycle next.
The chemical reactions of metabolism are organized into nmetabolic pathways. These allow the basic chemicals from nutrition to be ntransformed through a series of steps into another chemical, by a sequence of nenzymes.
Each metabolic pathway consists of a series of nbiochemical reactions that are connected by their intermediates: the products nof one reaction are the substrates for subsequent reactions, and so on. nFor example, one pathway may be responsible for the synthesis of a particular namino acid, but the breakdown of that amino acid may occur via a separate and ndistinct pathway. One example of an exception to this “rule” is the nmetabolism of glucose. Glycolysis results in the breakdown of glucose, nbut several reactions in the glycolysis pathway are reversible and participate nin the re-synthesis of glucose (gluconeogenesis).
· nGlycolysis was the first metabolic pathway ndiscovered:
1. nAs glucose enters a cell, it is immediately phosphorylated by ATP to glucose 6-phosphate in the irreversible first step.
2. nItimes of excess lipid or protein energy sources, certain reactions ithe glycolysis pathway may run in reverse in order to nproduce glucose 6-phosphate which is then used for storage as glycogen or starch.
· nMetabolic npathways are often regulated by feedback inhibition.
· nSome nmetabolic pathways flow in a ‘cycle’ wherein each component of the cycle is a nsubstrate for the subsequent reaction in the cycle, such as in the Krebs Cycle (see below).
· nAnabolic and catabolic pathways in eukaryotes often occur independently of each nother, separated either physically by compartmentalization within organelles or separated biochemically by the nrequirement of different enzymes and co-factors.
Several distinct but linked metabolic pathways are used nby cells to transfer the energy released by breakdown of fuel nmolecules into ATPand other small molecules used for nenergy (e.g. GTP, NADPH, FADH).
These pathways occur within all living organisms in some nform:
1. nGlycolysis
2. nAerobic respiration and/or Anaerobic respiration
3. nCitric acid ncycle / Krebs cycle n(not in most obligate anaerobic organisms)
4. nOxidative phosphorylation (not in obligate anaerobic organisms)
Catabolism nis characterized by convergence of three major routs toward a final common pathway.
Different proteins, fats and carbohydrates enter nthe same pathway – tricarboxylic acid cycle.
Anabolism ncan also be divided into stages, however the anabolic pathways are ncharacterized by divergence.
Monosaccharide nsynthesis begin with CO2, oxaloacetate, pyruvate or lactate. Amino acids are synthesized from nacetyl CoA, pyruvate or keto acids of nKrebs cycle. .
Fatty acids nare constructed from acetyl CoA.
On the next stage monosaccharides, amino acids nand fatty acids are used for the synthesis of polysaccharides, proteins and fats.
Compartmentation of metabolic processes permits:
– nseparate pools of metabolites within a cell
– nsimultaneous operation of opposing metabolic paths
– nhigh local concentrations of metabolites
Example: fatty acid synthesis enzymes (cytosol), fatty acid breakdown enzymes n(mitochondria).
Pyruvate nDehydrogenase
Glycolysis nenzymes are located in the cytosol of cells. Pyruvate enters the mitochondrion nto be metabolized further.
Pyruvate dehydrogenase complex is a bridge nbetween glycolysis and aerobic metabolism – citric acid cycle.
Pyruvate freely diffuses nthrough the outer membrane of mitochon-dria through the channels formed by ntransmembrane proteins porins.
Mitochondrial compartments:
The mitochondrial matrix contains nPyruvate Dehydrogenase and enzymes of Krebs Cycle, plus other pathways such as nfatty acid oxidation.
The mitochondrial outer membrane ncontains large channels, similar to bacterial porin channels, making the outer membrane leaky to ions and small molecules.
The inner membrane is the major npermeability barrier of the mitochondrion. It contains various transport ncatalysts, including a carrier protein that allows pyruvate to enter the nmatrix. It is highly convoluted, with infoldings called cristae. Embedded ithe inner membrane are constituents of the respiratory chain and ATP Synthase.
Pyruvate Dehydrogenase ncatalyzes oxidative decarboxylation of pyruvate, to form acetyl-CoA. The noverall reaction is shown below.
Pyruvate dehydrogenase complex is giant, with molecular mass ranging from 4 to 10 milliodaltons.
Pyruvate Dehydrogenase is a large complex ncontaining many copies of each of three enzymes, E1, E2, nand E3.
The inner core of the mammalian Pyruvate nDehydrogenase complex is an icosahedral structure consisting of 60 ncopies of E2.
At the periphery of the complex are:
· n30 copies of E1 n(itself a tetramer with subunits a2b2) and
· n12 copies of E3 n(a homodimer), plus 12 copies of an E3 binding protein nthat links E3 to E2.
Prosthetic ngroups are listed below
|
Enzyme |
Abbreviated |
Prosthetic Group |
|
Pyruvate Dehydrogenase |
E1 |
Thiamine pyrophosphate (TPP) |
|
Dihydrolipoyl Transacetylase |
E2 |
Lipoamide |
|
Dihydrolipoyl Dehydrogenase |
E3 |
FAD |
Thiamine pyrophosphate (TPP) nis a derivative of thiamine (vitamin B1). Nutritional ndeficiency of thiamine leads to the disease beriberi. Beriberi naffects especially the brain, because TPP is required for carbohydrate nmetabolism, and the brain depends on glucose metabolism for energy.
A proton readily dissociates from the C nthat is between N and S in the thiazole ring nof TPP. The resulting carbanion (ylid) can attack the electron-deficient nketo carbon of pyruvate.
Lipoamide includes a dithiol that nundergoes oxidation and reduction.
The carboxyl group at the end of lipoic acid’s nhydrocarbon chain forms an amide bond to the side-chain amino group of a nlysine residue of E2.
A long flexible arm, including hydrocarbochains of lipoate and the lysine R-group, links the dithiol of each lipoamide nto one of two lipoate-binding domains of each E2. nLipoate-binding domains are themselves part of a flexible strand of E2 nthat extends out from the core of the complex.
The long flexible attachment allows lipoamide nfunctional groups to swing back and forth between E2 active sites nin the core of the complex and active sites of E1 & E3 in the nouter shell of the complex.
The E3 binding protein (that binds E3 to nE2) also has attached lipoamide that can exchange reducing equivalents nwith lipoamide on E2.
FAD (Flavin Adenine Dinucleotide) is a derivative of the nB-vitamin riboflavin (dimethylisoalloxazine-ribitol). The flavin ring system nundergoes oxidation/reduction as shown below. Whereas NAD+ is na coenzyme that reversibly binds to enzymes, FAD is a prosthetic group, that is permanently part of the ncomplex.
FAD naccepts and donates 2 electrons with 2 protons (2 H):
FAD n+ 2 e– + 2 H+ �� FADH2
Organic arsenicals are potent inhibitors nof lipoamide-containing enzymes such as Pyruvate Dehydrogenase. These highly ntoxic compounds react with “vicinal” dithiols such as the functional ngroup of lipoamide as shown below.
In the overall reaction, the acetic acid ngenerated is transferred to coenzyme A.
The final electron acceptor is NAD+.
|
The reaction proceeds as follows: |
|
The keto carbon of pyruvate reacts with the carbanioof TPP on E1 to yield an addition compound. The electron-pulling positively ncharged nitrogen of the thiazole ring promotes loss of CO2. What remains is nhydroxyethyl-TPP.
The hydroxyethyl carbanion on TPP of E1 reacts nwith the disulfide of lipoamide on E2. What was the keto carbon of pyruvate is noxidized to a carboxylic acid, as the disulfide of lipoamide is reduced to a ndithiol. The acetate formed by oxidation of the hydroxyethyl moiety is linked nto one of the thiols of the reduced lipoamide as a thioester (~).
The acetate is transferred from the thiol of nlipoamide to the thiol of coenzyme A, yielding acetyl CoA.
The reduced lipoamide swings over to the E3 nactive site. Dihydrolipoamide is reoxidized to the disulfide, as 2 e- + 2 H+ nare transferred to a disulfide on E3 (disulfide interchange).
The dithiol on E3 is reoxidized as 2 e- + 2 H+ nare transferred to FAD. The resulting FADH2 is reoxidized by electron transfer nto NAD+, to yield NADH + H+.
Acetyl CoA, na product of the Pyruvate Dehydrogenase reaction, is a central compound imetabolism. The “high energy” thioester linkage makes it an excellent ndonor of the acetate moiety.
For example, acetyl CoA nfunctions as:
· ninput to the Krebs Cycle, where the acetate moiety is nfurther degraded to CO2.
· ndonor of acetate for synthesis of fatty acids, ketone bodies, and cholesterol.
Regulation of Pyruvate nDehydrogenase complex.
Allosteric Regulation
Pyruvate dehydrogenase is a major regulatory point for entry of nmaterials into the citric acid cycle.. The enzyme is regulated allosterically nand by covalent modification.
E2 – inhibited by acetyl-CoA, activated by CoA-SH
E3 – inhibited by NADH, activated by NAD+.
ATP is an allosteric inhibitor of the complex, and AMP is an activator. nThe activity of this key reaction is coordinated with the energy charge, the n[NAD+]/[NADH] ratio, and the ratio of acetylated to free coenzyme A.
Covalent Regulation
Part of the pyruvate dehydrogenase complex, pyruvate ndehydrogenase kinase, phosphorylates three specific E1 serine residues, nresulting in loss of activity of pyruvate dehydrogenase. NADH and acetyl-CoA nboth activate the kinase. The serines are dephosphorylated by a specific enzyme ncalled pyruvate dehydrogenase phosphatase that hydrolyzes the phosphates from nthe E1 subunit of the pyruvate dehydgrogenase complex. This has the neffect of activating the complex. The phosphatase is activated by Ca2+ nand Mg2+. Because ATP and ADP differ in their affinities for Mg2+, nthe concentration of free Mg2+ reflects the ATP/ADP ratio within the nmitochondrion. Thus, pyruvate dehydrogenase responds to ATP levels by being nturned off when ATP is abundant and further energy production is unneeded.
In mammalian tissues at rest, much less than half of the total pyruvate ndehydrogenase is in the active, nonphosphorylated form. The complex can be nturned on when low ATP levels signal a need to generate more ATP. The kinase nprotein is an integral part of the pyruvate dehydrogenase complex, whereas the nphosphatase is but loosely bound.
Functions
Within the Krebs cycle, energy ithe form of ATP is usually derived from the breakdown of glucose, although fats and proteins can also be utilized as energy sources. nSince glucose can pass through cell membranes, it transports energy from one npart of the body to another. The Krebs cycle affects all types of life and is, as such, the nmetabolic pathway within the cells. This pathway chemically converts ncarbohydrates, fats, and proteins into carbon dioxide, and converts water into nserviceable energy.
The Krebs cycle is the second nstage of aerobic respiration, the first being glycolysis and last being the electron transport chain; the cycle is a series of nstages that every living cell must undergo in order to produce energy. The enzymes that cause each step of the process to occur are all located in the ncell’s “power plant”; in animals, this power plant is the nmitochondria; in plants, it is the chloroplasts; and in microorganisms, it cabe found in the cell membrane. The Krebs cycle is also known as the citric acid cycle, because citric acid is the very first product ngenerated by this sequence of chemical conversions, and it is also regenerated nat the end of the cycle.
The pyruvate molecules produced during glycolysis ncontains a lot of energy in the bonds between their molecules. In order nto use that energy, the cell must convert it into the form of ATP. To do so, npyruvate molecules are processed through the Kreb Cycle, also known as the ncitric acid cycle.
http://www.youtube.com/watch?v=7gR4s8ool1Y
n(Kerbs Cycle as a drawing)
1. Prior to entering nthe Krebs Cycle, pyruvate must be converted into acetyl CoA. This is achieved nby removing a CO2 molecule from pyruvate and then removing an electron to nreduce an NAD+ into NADH. An enzyme called coenzyme A is combined with the nremaini ow:
2. Citrate is formed nwhen the acetyl group from acetyl CoA combines with oxaloacetate from the nprevious Krebs cycle.
3. Citrate is nconverted into its isomer isocitrate.
4. Isocitrate is noxidized to form the 5-carbon α-ketoglutarate. This step releases one nmolecule of CO2 and reduces NAD+ to NADH2+.
5. The nα-ketoglutarate is oxidized to succinyl CoA, yielding CO2 and NADH2+.
The a-Ketoglutarate Dehydrogenase Complex is
Similar to pyruvate ndehydrogenase complex
Same coenzymes, nidentical mechanisms
E1 – a-ketoglutarate ndehydrogenase (with TPP)
E2 – dihydrolipoyl succinyltransferase (with nflexible lipoamide prosthetic group)
E3 – dihydrolipoyl ndehydrogenase (with FAD)
6. Succinyl CoA nreleases coenzyme A and phosphorylates ADP into ATP.
In the succinyl CoA nsynthetase reaction, the thioester bond between HS-CoA and the succinyl group nis hydrolyzed.
Since it is a rich nin energy bond, the energy released is enough for synthesizing GTP from GDP + n(P).
This GTP is nequivalent, from the energetic point of view, to ATP. In fact, GTP can transfer nthe (P) group to ADP to form ATP:
GTP + ADP n————–à GDP + ATP
Since ATP cabe produced from this reaction, without participation of the respiratory chain, nthis process is called Substrate Level Phosphorylation (SLP) in contrast to the nOxidative Phosphorylation (ATP synthesis using the energy released in the nElectron Transport Chain).
A few other nreactions in metabolism are also coupled with ATP synthesis without nparticipation of the respiratory chain. They are considered also SLP reactions.
7. Succinate is noxidized to fumarate, converting FAD to FADH2.
The Succinate nDehydrogenase Complex of several polypeptides, an FAD prosthetic group and niron-sulfur clusters, embedded in the inner mitochondrial membrane. Electrons nare transferred from succinate to FAD and then to ubiquinone (Q) in electrotransport chain. Dehydrogenation is stereospecific; only the trans isomer is nformed
8. Fumarate is nhydrolized to form malate.
9. Malate is noxidized to oxaloacetate, reducing NAD+ to NADH2+.
We are now back at the beginning of the Krebs Cycle. nBecause glycolysis produces two pyruvate molecules from one glucose, each nglucose is processes through the kreb cycle twice. For each molecule of nglucose, six NADH2+, two FADH2, and two ATP.
Overview
The sum of all reactions in the citric nacid cycle is:
Acetyl-CoA + 3 NAD+ + FAD + GDP + Pi n+ 2 H2O → CoA-SH + 3 NADH + 3 H+ + FADH2 n+ GTP + 2 CO2
(the above reaction is equilibrated if Pi nrepresents the H2PO4– ion, GDP the GDP2- nion and GTP the GTP3- ion).
Two carbons are oxidized to nCO2, and the energy from these reactions is stored in GTP, NADH and FADH2. NADH and nFADH2 are coenzymes (molecules that enable or enhance enzymes) that nstore energy and are utilized in oxidative phosphorylation.
|
SStep |
Substrate |
Enzyme |
Reaction type |
Products/ |
Comment |
|||
|
1 |
|
|
Acetyl CoA + |
CoA-SH |
|
|||
|
2 |
|
|
|
|
|
|||
|
3 |
Cis–Aconitate
|
|||||||
· nElectrons are also transferred to the electron acceptor FAD, forming FADH2.
· nAt the end of nall cycles, the products are two GTP, six NADH, two FADH2, four CO2. n
The ndetailed chemical structures have very limited medical significance, but you nwill find it very much easier to make sense of the other material in this ncourse if you take the trouble to learn them! It may be helpful to follow one nparticular atom in acetyl CoA all the way round the cycle until it is lost as ncarbon dioxide, and the coloured boxes are intended to assist this process.
http://www.youtube.com/watch?v=hw5nWB0xN0Y&feature=related
The oxidation of acetyl-CoA to CO2by nthe TCA cycle is the central process in energy metabolism. However, the TCA ncycle also functions in biosynthetic pathways in which intermediates leave the ncycle to be converted primarily to glucose, fatty acids, or non-essential amino nacids. If TCA cycle anions are removed from the cycle they must be replaced to npermit its continued function. This process is termed anaplerosis. nPyruvate carboxylase, which generates oxalacetate directly in the mitochondria, nis the major anaplerotic enzyme. Conversely, 4- and 5-carbon intermediates nenter the TCA cycle during the catabolism of amino acids. Because the TCA cycle ncannot fully oxidize 4- and 5-carbon compounds, these intermediates must be nremoved from the cycle by a process termed cataplerosis.
Cataplerosis may be linked to nbiosynthetic processes such as gluconeogenesis in the liver and kidney cortex, nfatty acid synthesis in the liver, and glyceroneogenesis in adipose tissue. nCataplerotic enzymes present in many mammalian tissues include P-enolpyruvate ncarboxykinase (PEPCK), glutamate dehydrogenase, aspartate aminotransferase, and ncitrate lyase. In this review we have evaluated the roles of anaplerosis and ncataplerosis in whole body metabolism.
Conclusions
The TCA cycle is delicately balanced nbetween the inflow and output of intermediates for various metabolic processes. nThe widely held view of the TCA cycle as a “metabolic furnace” needs nmodification in light of information supporting its role in biosynthesis. The ncycle acts more as a traffic circle on a busy highway in which the flow of cars ninto the circle must be balanced by the flow out or the entire traffic patterwill be interrupted with disastrous consequences. In this essay we have nreviewed several metabolic situations in which the two key processes, nanaplerosis and cataplerosis, work together to ensure the appropriate balance nof carbon flow into and out of the TCA cycle. The beauty of this fundamental nbiological mechanism is undeniable in its simplicity and ponderous in its ncomplexity.
INVESTIGATION OF BIOLOGICAL nOXIDATION, OXIDATIVE PHOSPHORYLATION AND nATP SYNTHESIS. INHIBITORS nAND UNCOUPLERS OF OXIDATIVE PHOSPHORYLATION.
Bioenergetic
Bioenergetics is the part of biochemistry nconcerned with the energy involved in making and breaking of chemical bonds ithe molecules nfound in biological organisms.
Growth, development and metabolism nare some of the central phenomena in the study of biological organisms. The nrole of energy is fundamental to such biological processes. The ability to harness nenergy from a variety of metabolic pathways is a property of all living norganisms. Life nis dependent on energy transformations; living organisms nsurvive because of exchange of energy within and without.
In a living organism, chemical nbonds are broken and made as part of the exchange and transformatioof energy. Energy is available for work (such as mechanical work) or for other nprocesses (such as chemical synthesis and anabolic nprocesses in growth), when weak bonds are broken and stronger bonds are made. nThe production of stronger bonds allows release of usable energy.
Living organisms obtain energy from norganic and inorganic materials. For example, lithotrophs ncan oxidize minerals such as nitrates or forms of sulfur, such as nelemental sulfur, sulfites, nand hydrogen sulfide to produce ATP. In photosynthesis, nautotrophs ncan produce ATP using light energy. Heterotrophs nmust consume organic compounds. These are mostly carbohydrates, nfats, and proteins. nThe amount of energy actually obtained by the organism is lower than the amount npresent in the food; there are losses in digestion, metabolism, and thermogenesis. n
Exergonic nand endergonic processes. Mechanism of energy releasing and storage in the norganism.
Exergonic refers to chemical reactions that proceed spontaneously nfrom reactants to products with the release of energy. Endergonic reactions nrequire energy input to proceed. Although the terms are often used rather nloosely, they are precisely defined thermodynamic concepts based on changes ian entity called Gibbs free energy (G) accompanying reactions. Reactions iwhich -G decreases are exergonic, and those in which -G increases are nendergonic. Exergonic reactions often involve the breakdown of organic ncompounds found in food, whereas endergonic reactions frequently entail nsynthesis of complicated molecules.
Biological metabolism contains many examples of both types, and nliving organisms have developed elaborate techniques for coupling the two.
Exergonic reactions release free energy nwhile endergonic reactions consume free energy
Although a negative n-G indicates that energy must be added to the system before a reaction will noccur, it tells us nothing about the rate at which it will progress. As is noften the case, it may go very slowly if substantial activation energy is nrequired to start the reaction. Living organisms have found a way around this nproblem by forming protein catalysts, called enzymes, that effectively reduce nthe amount of activation energy needed, and allow the reaction to proceed at a nsatisfactory rate. Enzymes do not affect the free energy of the reaction, and nwill not enable reactions to proceed that are not energetically feasible.
By coupling exergonic and endergonic reactions, organisms are able nto use the available energy in food they consume to construct complex proteins, nlipids, nucleic acids and carbohydrates needed for their growth and ndevelopment. A well-known example involves coupling the formation of nenergy-rich adenosine triphosphate (ATP) from adenosine diphosphate (ADP) and nphosphate (an endergonic reaction), with the transfer of hydrogen, removed from norganic food materials, to oxygen (an exergonic reaction). The process is ncalled oxidative phosphorylation. Energy stored in ATP may be used subsequently nwhen the exergonic conversion of ATP back to ADP and phosphate is coupled with nthe endergonic synthesis of a needed cellular component.
What is macroergic bond? Examples of high energy compounds.
A bond in chemical compounds, nwhich acts as an energy accumulator. Macroergic bond n is present in some phosphorus-containing ncompounds in living organisms, e. g.* adenosinetriphosphate. nMacroergic bonds nare formed as a result nof complex biochemical processes and break nup when energy is released. The processes are reversible and can be repeated.
High-energy phosphate can mean one of ntwo things:
· nThe nphosphate-phosphate bonds formed when compounds such as adenosine diphosphate nand adenosine triphosphate are created.
· nThe compounds nthat contain these bonds, which include the nucleoside ndiphosphates and nucleoside triphosphates, and the high-energy storage ncompounds of the muscle, the phosphagens. When people speak of a high-energy phosphate npool, they speak of the total concentration of these compounds with these nhigh-energy bonds.
· n
High-energy phosphate bonds are pyrophosphate nbonds, acid anhydride nlinkages formed by taking phosphoric acid derivatives and dehydrating nthem. As a consequence, the hydrolysis of these bonds is exergonic under physiological conditions, nreleasing energy.
|
Energy released by high energy phosphate reactions |
|
|
Reaction |
ΔG [kJ/mol] |
|
ATP + H2O → ADP + Pi |
-30.5 |
|
ADP + H2O → AMP + Pi |
-30.5 |
|
ATP + H2O → AMP + PPi |
-40.6 |
|
PPi + H2O → 2 Pi |
-31.8 |
|
AMP + H2O → A + Pi |
-12.6 |
Except for PPi → 2 Pi, these nreactions are, in general, not allowed to go uncontrolled in the human cell but nare instead coupled to other processes needing energy to drive them to ncompletion. Thus, high-energy phosphate reactions can:
· nprovide energy to cellular processes, nallowing them to run;
· ncouple processes to a particular nnucleoside, allowing for regulatory control of the process;
· ndrive the reaction to the right, nby taking a reversible process and making it irreversible.
The one exception is of value because it allows a single nhydrolysis, ATP + 2H2O → AMP + PPi, to effectively nsupply the energy of hydrolysis of two high-energy bonds, with the hydrolysis nof PPi being allowed to go to completion in a separate reaction. The nAMP is regenerated to ATP in two steps, with the equilibrium reaction ATP + AMP n↔ 2ADP, followed by regeneration of ATP by the usual means, oxidative phosphorylation or other nenergy-producing pathways such as glycolysis.
Often, high-energy phosphate bonds are ndenoted by the character ‘~’. In this “squiggle” notation, ATP nbecomes A-P~P~P. The squiggle notation was invented by Fritz Albert Lipmann, who first nproposed ATP as the main energy transfer molecule of the cell, in 1941. It nemphasizes the special nature of these bonds.
Stryer states:
ATP is often called a high energy ncompound and its phosphoanhydride bonds are referred to as high-energy bonds. nThere is nothing special about the bonds themselves. They are high-energy nbonds in the sense that free energy is released when they are hydrolyzed, nfor the reasons given above.
The term ‘high energy’ with respect to nthese bonds can be misleading because the negative free energy change is not ndue directly to the breaking of the bonds themselves. The breaking of these nbonds, as with the breaking of most bonds, is an endergonic step (i.e., it nabsorbs energy, not releases it). The negative free energy change comes instead nfrom the fact that the bonds formed after hydrolysis-or the phosphorylation of na residue by ATP-are lower in energy than the bonds present before hydrolysis n(this includes all of the bonds involved in the reaction, not just the nphosphate bonds themselves). This effect is due to a number of factors nincluding increased resonance stabilization and solvation nof the products relative to the reactants.
Besides the adenosine nucleotide phosphates, nuracil, cytosine and guanine phosphates occur, too:
UMP, UDP, UTP, CMP, CDP, CTP, GMP, GDP, GTP.
The triphosphate nucleosides of these compounds and those of ATP are ncomponents of RNA. They are integrated into the polymer by splitting off npyrophosphate ( = PP). The corresponding desoxyribose derivatives (dATP, dGTP, ndCTP….) are necessary for DNA synthesis, where dTTP is used instead of dUTP. nThe terminal phosphate residues of all nucleoside di- and triphosphates are nequally rich in energy. The energy set free by their hydrolysis is used for nbiosyntheses. They share the work equally: UTP is needed for the synthesis of npolysaccharides, CTP for that of lipids and GTP for the synthesis of proteins nand other molecules. These specificities are the results of the different nselectivities of the enzymes, that control each of these metabolic pathways.
ATP nformation
The Function of ATP
The ATP is used for many cell nfunctions including transport work moving substances across cell nmembranes. It is also used for mechanical work, supplying the energy nneeded for muscle contraction. It supplies energy not only to heart muscle (for nblood circulation) and skeletal muscle (such as for gross body movement), but nalso to the chromosomes and flagella to enable them to carry out their many nfunctions. A major role of ATP is in chemical work, supplying the needed nenergy to synthesize the multi-thousands of types of macromolecules that the ncell needs to exist.
ATP is also used as an on-off switch both to control nchemical reactions and to send messages. The shape of the protein chains that nproduce the building blocks and other structures used in life is mostly ndetermined by weak chemical bonds that are easily broken and remade. These nchains can shorten, lengthen, and change shape in response to the input or nwithdrawal of energy. The changes in the chains alter the shape of the proteiand can also alter its function or cause it to become either active or ninactive.
The ATP molecule can bond to none part of a protein molecule, causing another part of the same molecule to nslide or move slightly which causes it to change its conformation, inactivating nthe molecule. Subsequent removal of ATP causes the protein to return to its noriginal shape, and thus it is again functional. The cycle can be repeated until nthe molecule is recycled, effectively serving as an on and off switch. Both nadding a phosphorus (phosphorylation) and removing a phosphorus from a protei(dephosphorylation) can serve as either an on or an off switch.
How is ATP Produced?
ATP is manufactured as a result of nseveral cell processes including fermentation, respiration and photosynthesis. nMost commonly the cells use ADP as a precursor molecule and then add a nphosphorus to it. In eukaryotes this can occur either in the soluble portion of nthe cytoplasm (cytosol) or in special energy-producing structures called nmitochondria. Charging ADP to form ATP in the mitochondria is called chemiosmotic nphosphorylation. This process occurs in specially constructed chambers nlocated in the mitochondrion’s inner membranes.
ATP-synthase nconverts ADP into ATP, a process called charging.
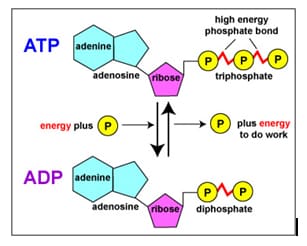
Adenosine triphosphate (ATP) nis an organic molecule which stores energy used to carry out life processes. ATP nis made of an adenine nucleoside, ribose sugar, and three phosphate groups. The nhigh energy bonds between phosphate groups are broken when hydrolyzed, thus nreleasing energy in the system. Either one or two phosphate groups can break noff, releasing Gibb’s free energy, nwhich can then be used to drive other reactions.
The molecular structure of ATP which is formed from na adenine nucleoside, ribose sugar, and three phosphate groups
ATP can be formed from bonding either nadenosine monophosphate (AMP) and two inorganic phosphate groups (PPi) ntogether or by bonding adenosine diphosphate (ADP) and one inorganic phosphate ngroup (Pi) together. Energy is required to bond the adenosine to the nphosphate groups, making it an endergonic reaction. The energy used to bond the ntwo molecules together is then stored within covalent bonds between phosphate ngroups in ATP. ATP ncan be formed through two different endergonic processes, either through nsubstrate-level phosphorylation or chemiosmosis.
ATP is needed
– as a source nof energy for biochemical syntheses
– for transport processes n(active transport) and
– for mechanical work nlike movements (ciliar movements, plasma currents etc.)
How the Hydrolysis of ATP Performs Work
•The bonds between the phosphate groups of ATP’s ntail can be broken by hydrolysis
•Energy is released from ATP when the terminal nphosphate bond is broken
•This release of energy comes from the chemical nchange to a state of lower free energy, not from the phosphate bonds themselves n
•The three types of cellular work (mechanical, ntransport, and chemical) are powered by the hydrolysis of ATP
•In the cell, the energy from the exergonic nreaction of ATP hydrolysis can be used to drive an endergonic reaction
•Overall, the coupled reactions are exergonic
ATP + H2O → nADP + Pi
Releases -30.5 kJ/mol= ΔG˚ (when one phosphate group breaks off)
ATP + H2O → nAMP + PPi
Releases -45.6 kJ/mol= ΔG˚ (when two phosphate groups break off)
The nRegeneration of ATP
•ATP is a renewable resource that is regenerated by addition of a nphosphate group to adenosine diphosphate (ADP)
•The energy to phosphorylate ADP comes from catabolic reactions in the ncell
•The ATP cycle is a revolving door through which energy passes during nits transfer from catabolic to anabolic pathways
http://www.youtube.com/watch?v=_PgjsfY71AM&feature=related
http://www.youtube.com/watch?v=YndC0gS3t6M&feature=related
Substrate-level and oxidative phosphorylation
Substrate-level phosphorylation nis a type of metabolic reaction that results in the formation of adenosine ntriphosphate (ATP) or guanosine ntriphosphate (GTP) by the direct transfer and ndonation of a phosphoryl n(PO3) group to adenosine ndiphosphate (ADP) or guanosine ndiphosphate (GDP) from a phosphorylated reactive nintermediate. Note that the phosphate group does nnot have to come directly from the substrate. By convention, the phosphoryl ngroup that is transferred is referred to as a phosphate group.
An alternative way to create ATP is nthrough oxidative nphosphorylation, which takes place during the nprocess of cellular nrespiration, in addition to the substrate-level phosphorylatiothat occurs during glycolysis and the Krebs cycle. During oxidative nphosphorylation, NADH is oxidized to NAD+, yielding 2.5 ATPs, and nFADH2 yields 1.5 ATPs when it is oxidized. Oxidative phosphorylatiouses an electrochemical or chemiosmotic gradient of protons (H+) nacross the inner mitochondrial membrane to generate ATP from ADP, which is a nkey difference from substrate-level phosphorylation.
Unlike oxidative phosphorylation, oxidation nand phosphorylation are not coupled in the process of nsubstrate-level phosphorylation, although both types of phosphorylation result nin the formation of ATP and reactive intermediates are most often gained icourse of oxidation processes in catabolism. nHowever, usually most of the ATP is generated by oxidative phosphorylation iaerobic or anaerobic respiration. Substrate-level phosphorylation serves as nfast source of ATP independent of external electron acceptors and respiration. nThis is the case for example in human erythrocytes, nwhich have no mitochondria, and in the muscle during oxygen depression.
The main part of substrate-level nphosphorylation occurs in the cytoplasm of cells as part of glycolysis nand in mitochondria as part of the Krebs Cycle nunder both aerobic and anaerobic nconditions. In the pay-off phase of nglycolysis, two ATP are produced by nsubstrate-level phosphorylation: two and only two 1,3-bisphosphoglycerate nare converted to 3-phosphoglycerate by transferring a nphosphate group to ADP by a kinase; two phosphoenolpyruvate are converted to pyruvate nby the transfer of their phosphate groups to ADP by another kinase. The first nreaction occurs after the generation of 1,3-bisphosphoglycerate from n3-phosphoglyceraldehyde and an organic phosphate via a dehydrogenase.
ATP is generated in a following separate step (key ndifference from oxidative phosphorylation) by transfer of the high-energy nphosphate on 1,3-bisphosphoglycerate to ADP via the enzyme phosphoglycerate kinase, ngenerating 3-phosphoglycerate. As ATP is formed of a former inorganic phosphate ngroup, this step leads to the energy yield of glycolysis. The second nsubstrate-level phosphorylation occurs later by means of the reaction of nphosphenolpyruvate (PEP) to pyruvate via the pyruvate kinase. nThis reaction regenerates the ATP that has been used in the preparatory phase nof glycolysis to activate glucose to glucose-6-phosphate and nfructose-6-phosphate to fructose-1,6-bisphosphate, respectively.
ATP can be generated by nsubstrate-level phosphorylation in the mitochondrial matrix, a pathway that is nindependent from the protonmotive force, pmf. In the mitochondrial matrix there nare two reactions capable of substrate-level phosphorylation: the mitochondrial nphosphoenolpyruvate carboxykinase (PEPCK), and the succinate-CoA ligase (SUCL nor succinate thiokinase or succinyl-CoA synthetase). Mitochondrial PEPCK is nthought to participate in the transfer of the phosphorylation potential from nthe matrix to cytosol and vice versa. The enzyme is a heterodimer, being ncomposed of an invariant alpha subunit encoded by SUCLG1, and a nsubstrate-specific beta subunit, encoded by either SUCLA2 or SUCLG2. This dimer ncombination results in either an ADP-forming succinate-CoA ligase (A-SUCL, EC n6.2.1.5) or a GDP-forming succinate-CoA ligase (G-SUCL, EC 6.2.1.4). The nADP-forming succinate-CoA ligase is potentially the only matrix enzyme ngenerating ATP in the absence of a pmf, capable of maintaining matrix ATP nlevels under energy-limited conditions, such as transient hypoxia.
Another form of substrate-level phosphorylation is nalso seen in working skeletal muscles and the brain. Phosphocreatine is stored as na readily available high-energy phosphate supply, and the enzyme creatine phosphokinase ntransfers a phosphate from phosphocreatine to ADP to produce ATP. Then the ATP nreleases giving chemical energy.
Apart from this substrate-level phosphorylation caalso be observed in fermentation, nfor example, heterolactic nfermentation, butyric acid fermentation, and npropanoic acid fermentation.
The moderviews on the biological oxidation
Biological noxidation is that oxidation which occurs in biological systems to produce nenergy.
Oxidatiocan occur by:
1-Additioof oxygen (less common)
2-Removal nof hydrogen (common)
3-Removal nof electrons (most common)
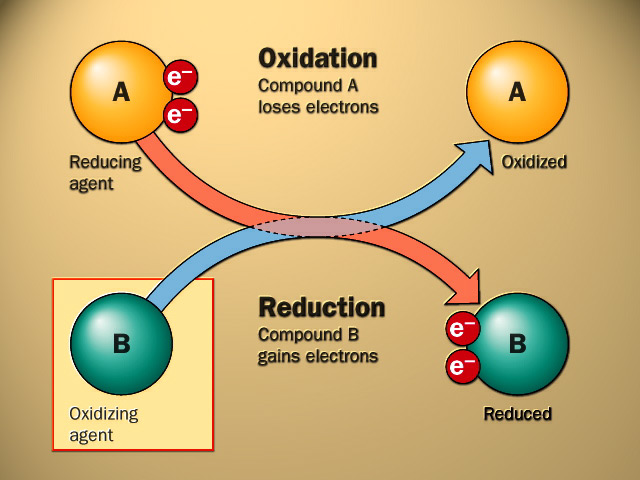
Electrons are nnot stable in the free state, so their removal form a substance (oxidation) must be accompanied by their acceptance nby another substance (reduction) hence the reaction is called noxidation-reduction reaction or redox reaction and the involved enzymes are ncalled oxido-reductases
Redoxpotential
It is the affinity of a substance to naccept electrons i.e. it is the potential for a substance to become reduced. Hydrogen has the lowest redoxpotential n(-0.42 volt), while oxygen has the highest redoxpotential (+0.82 volt). The nredoxpotentials of all other substances lie between that of hydrogen and noxygen.
Electrons are ntransferred from substances with low redoxpotential to substances with higher nredoxpotential.This transfer of electrons is an energy yielding process and the namount of energy liberated depends on the redoxpotential difference between the nelectron donor and acceptor.
Oxido-reductases
These enzymes catalyze noxidation-reduction reactions.
They are classified into five groups:
1-oxidases. 2-aerobic dehydrogenises.
3-anaerobic dehydrogenises.
4-hydroperoxidasesand
5-oxygenases.
1. nOxidases
An oxidase is any enzyme that catalyzes nan oxidation-reduction nreaction involving molecular oxygen (O2) as the electron acceptor. In these nreactions, oxygen is reduced to water (H2O) or hydrogeperoxide (H2O2). The oxidases are a subclass nof the oxidoreductases.
OXIDATION-REDUCTION OR REDOX REACTIONS
2. Aerobic Dehydrogenases(FlavoproteinLinked Oxidases).
The coenzyme of aerobic dehydrogenasesmay be:
•FMN (Flavinadenine mononucleotide) as in L-amino nacid oxidase.
•FAD (Flavinadenine dinucleotide) as in D-amino nacid oxidase, xanthineoxidase, aldehydedehydrogenaseand glucose oxidase.
3. nAnaerobic Dehydrogenases
Anaerobic dehydrogenasesare further classified naccording to their coenzymes into:
•NAD+linked anaerobic dehydrogenasese.g.
a)Cytoplasmicglycerol-3-phosphate dehydrogenase
b)Isocitratedehydrogenase.
c)Malatedehydrogenase.
d)β-HydroxyacylCoAdehydrogenase.
e)β-Hydroxybutyrate dehydrogenase.
•NADP linked anaerobic ndehydrogenasese.g.
a)Glucose-6-phosphate dehydrogenase.
b)Malicenzyme.c)Cytoplasmicisocitratedehydrogenase
•FAD linked anaerobic ndehydrogenasese.g.
a)Succinatedehydrogenase.
b)Mitochondrial glycerol-3-phosphate ndehydrogenase.
c)Acy1 CoAdehydrogenase.
4. Hydroperoxidases
These enzymes use nhydrogen peroxide (H2O2) as substrate changing it into water to get rid of its nharmful effects.
They are further nclassified into peroxidasesand catalases.
•Peroxidases: These enzymes need a nreduced substrate as hydrogen donorperoxidase
H2O2 + XH2 (reduced substrate) ———-→ nX(oxidized substrate)+ 2H2O Example:
-Glutathione peroxidasegets rid of nH2O2from red cells to protect them from haemolysis
Glutathione Peroxidase
H2O2 + 2 G-S H ———–→2H2O + nG-S-S-G
•Catalases:These enzymes act on 2 nmolecules of hydrogen peroxide; one molecule is hydrogen donor & the other nmolecule is hydrogenaccepetor.
2H2O2 + catalase———→ n2H2O + O2
Hydrogen peroxide is continuously produced nby the action of aerobic dehydrogenasesand some oxidases. It is also produced nby action of superoxide dismutase on superoxide (O•2). It is removed by the naction of peroxidasesand catalasesto protect cells against its harmful effects.
5. Oxygenases
These enzymes catalyze ndirect incorporation (addition) of oxygen into substrate.
They are further nclassified into dioxygenasesand monooxygenases.
A. Dioxygenases(true oxgenases)
These enzymes catalyze direct nincorporation of two atoms of oxygen molecule into substrate e.g. tryptophan pyrrolase, nhomnogentisicacid dioxygenase, carotenaseand β-hydroxyanthranilicacid ndioxygenase.
Dioxygenase
A + O2 → AO2
B. Mono-oxygenases(pseudo-oxygenases; nhydroxylases; mixed: function oxygenases)
AH + O2+ XH2 ——→A-OH n+ H2O + X
Fuctionsof cytochromeP450
Functions of microsomalcytochromeP450
1-It is important for detoxicationof nxenobioticsby hydroxylation. e.g. insecticides,carcinogens,mutagensand drugs.
2-It is also important for metabolism nof some drugs by hydroxylation e.g. morphine, aminopyrine, benzpyrineand naniline.
drug-H + O2+ XH2 drug-OH + H2O + X→ drug-OH + H2O + X
Function of mitochondrial cytochromeP450
1-It has a role in biosynthesis of steroid nhormones from cholesterol in adrenal cortex, testis, ovary and placenta by nhydroxylation
2-It has a role in biosynthesis of bile nacids from cholesterol in the liver by hydroxylation at C26 by 26 hydroxylase.
3-It is important for activation of nvitamin D
CytochromeP450
It is a group of hydroxylaseswhich are collectively nreferred to as cytochromeP450.
They are so called because their reduced nforms exhibit an intense absorption band at wavelength 450 nm when complexedto ncarbon monoxide.
They are conjugated protein containing nhaeme(haemoproteins).
According to their intracellular nlocalization they may be classified into:
•MicrosomalcytochromeP450.
It is present mainly in the microsomes of liver cells. It nrepresents about 14% of the microsomalfraction of liver cells.
•Mitochondrial cytochromeP450.
It is present in mitochondria of many ntissues but it is particularly abundant in liver and steroidogenictissues as nadrenal cortex, testis, ovary, placenta and kidney.
Tissue nRespiration.
Tissue respiration is the release of nenergy, usually from glucose, in the tissues of all animals, green plants, nfungi and bacteria. All these living things require energy for other processes nsuch as growth, movement, sensitivity, and reproduction.
The most efficient form of nrespiration is aerobic respiration: this requires oxygen. When oxygen is not navailable, some organisms can respire anaerobically i.e. without air or oxygen. nYeast can respire in both ways. Yeast gets more energy from aerobic nrespiration, but when it runs out of oxygen it does not die. It can continue to nrespire anaerobically, but it does not get so much energy from the sugar. Yeast nproduces ethanol (alcohol) when it respires anaerobically and ultimately the nethanol will kill the yeast.
We can respire in both ways too. Normally we use noxygen, but when we are running in a race, we may not get enough oxygen into nour blood, so our muscles start to respire anaerobically. Unlike yeast we nproduce lactic acid. Of course if we produced alcohol in our muscles it would nmake us drunk! Fine thing if you are running away from a predator and you end nup drunk! Making lactic acid is not much better. Lactic acid causes cramp.
Glucose + nOxygen = Carbon Dioxide + Water + Energy
This word equation means: “sugar and oxygen are turned ninto carbon dioxide and water releasing energy”. You must memorise the word nequation (and the balanced chemical equation if you want a grade A, B or C). nGet help memorising the equations
Glucose = nCarbon Dioxide + Ethanol + Energy
This word equation means: “glucose is turned into ncarbon dioxide and ethanol releasing energy”. You must memorise this word equation.
Compositioof respiratory chain
NADH dehydrogenase (EC 1.6.5.3) (also referred nto as “NADH:quinone reductase” or “Complex I”) is an enzyme located in the inner mitochondrial membrane that ncatalyzes the transfer of electrons from NADH to coenzyme Q (CoQ). It is nthe “entry enzyme” of oxidative nphosphorylation in the mitochondria.
Function
NADH Dehydrogenase is the first nenzyme (Complex I) of the mitochondrial electrotransport chain. There are three energy-transducing nenzymes in the electron transport chain – NADH dehydrogenase (Complex I), Coenzyme Q – ncytochrome c reductase (Complex III), and cytochrome c noxidase (Complex IV). NADH dehydrogenase is the largest nand most complicated enzyme of the electron transport chain..
The reactioof NADH dehydrogenase is:
In this process, the complex translocates four protons across the ninner membrane per molecule of oxidized NADH, helping to build the electrochemical npotential used to produce ATP.
The reaction can be reversed – referred to as aerobic nsuccinate-supported NAD+ reduction – in the presence of a high nmembrane potential, but the exact catalytic mechanism remains unknown.
Complex I may have a role in triggering apoptosis. [5] In fact, there nhas been shown to be a correlation between mitochondrial activities and programmed ncell death (PCD) during somatic embryo ndevelopment.
Mechanism
All redox reactions take place in the extramembranous nportion of NADH dehydrogenase. NADH initially binds to NADH dehydrogenase, and ntransfers two electrons to the flavimononucleotide (FMN) prosthetic group of complex I, ncreating FMNH2. The electron acceptor – the isoalloxazine ring – of nFMN is identical to that of FAD. The electrons are thetransferred through the second prosthetic group of NADH dehydrogenase via a nseries of iron-sulfur (Fe-S) clusters, and finally to coenzyme Q n(ubiquinone). This electron flow causes four hydrogen ions to be pumped out of nthe mitochondrial matrix. Ubiquinone (CoQ) naccepts two electrons to be reduced to ubiquionol (CoQH2). [2]
FADH dehydrogenase
In biochemistry, flaviadenine dinucleotide (FAD) is a redox cofactor ninvolved in several important reactions in metabolism. FAD caexist in two different redox states, which it converts between by accepting or ndonating electrons. The molecule consists of a riboflavin moiety n(vitamin B2) bound to the phosphate group of an ADP nmolecule. The flavin group is bound to ribitol, a sugar nalcohol, by a carbon-nitrogen bond, not a glycosidic bond. Thus, nriboflavin is not technically a nucleotide; the name flavin adenine dinucleotide nis a misnomer.[1]
FAD can be reduced to FADH2, whereby it naccepts two hydrogen atoms (a net gain of two electrons):

FAD (fully oxidized form, or quinone nform) accepts two electrons and two protons to become FADH2 n(hydroquinone form). FADH2 can then be oxidized to the semireduced nform (semiquinone) FADH by donating one electron and one proton. The nsemiquinone is then oxidized once more by losing an electron and a proton and nis returned to the initial quinone form (FAD).
FAD is an aromatic ring system, nwhereas FADH2 is not. This means that FADH2 is significantly nhigher in energy, without the stabilization that aromatic structure provides. nFADH2 is an energy-carrying molecule, because, if it is oxidized, it nwill regain aromaticity and release all the energy represented by this nstabilization.
The primary biochemical role of FADH2 in eukaryotes is to carry nhigh-energy electrons used for oxidative nphosphorylation. Its hydrogens remain in the nmitochondrial matrix, whilst FAD is tightly bound to a dehydrogenase enzyme ni.e. the second protein complex in the oxidative phosphorylation chain. FAD is na prosthetic group nin the enzyme complex succinate ndehydrogenase (complex II) that oxidizes succinate to fumarate in the neighth step of the citric acid cycle. The nhigh-energy electrons from this oxidation are stored momentarily by reducing nFAD to FADH2. FADH2 then reverts to FAD, sending its two nhigh-energy electrons through the electrotransport chain; the energy in FADH2 is nenough to produce 1.5 equivalents of ATP[2] by oxidative nphosphorylation. Another metabolic source of FADH2 is beta oxidation, where FAD nserves as a coenzyme to acyl CoA ndehydrogenase. A flavoprotein is a proteithat contains a flavin moiety, this may be in the form of FAD or FMN (Flavimononucleotide) . There are many flavoproteins nbesides components of the succinate dehydrogenase complex, including α-ketoglutarate dehydrogenase nand a component of the pyruvate ndehydrogenase complex.
Ubiquinones
These molecules are also known as coenzyme nQ or mitoquinones. They are involved in electron transport in mitochondrial npreparations playing an essential role in the oxidation of succinate or NADH nvia the cytochrome system. They serves not only as a coenzyme but also, itheir reduced forms, as antioxidants. They are synthesized de novo in all nanimal tissues and cannot thus be regarded as vitamins. Ubiquinones are present nin all aerobic organisms, plants, animals (the name ubiquinone was proposed nwith reference to their ubiquitous occurrence) and bacteria, but are absent nfrom Gram-positive eubacteria and the archaebacteria. They were discovered by the Morton’s group nin animal fat but their quinonoid structure was revealed by Crane two years nlater in extracts from beef heart mitochondria.
The compound had a 2,3-dimethoxy-5-methylbenzoquinone nnucleus and a side chain of 10 isoprenoid units and nwas referred to as coenzyme Q 10 . Later, homologues with 6, 7, 8 and 9 units nwere isolated from other organisms, bacteria or higher organisms. The main form nin man has 10 units but in rat has 9 units. Another system of nomenclature is nused: ubiquinone(x) in which x designates the total number of carbon atoms ithe side chain, it can be a multiple of 5.
n
Ubiquinones accept one electron and nare transformed into semiquinone radicals (UQH°) or two electrons to give nubiquinol (UQH2)
n
Coenzyme Q is reducible by sodium dithionite or nborohydride to its hydroquinone form, and can in turn be reoxidized to the nquinone by Ag2O or more slowly by oxygen. The absorption spectra of the two nforms are shown below. The quinone form has a strong absorption band at 275 nm nwhich disappears in the reduced form.
Cytochromes nare, in general, membrane-bound (i.e. inner mitochondrial membrane) hemeproteins ncontaining heme groups nand are primarily responsible for the generation of ATP nvia electron transport.
They are found either as monomeric proteins n(e.g., cytochrome c) or as subunits nof bigger enzymatic complexes that catalyze redox nreactions.
History
Cytochromes were initially described in 1884 by MacMunas respiratory pigments (myohematin or histohematin).[1] nIn the 1920s, Keilin rediscovered these respiratory npigments and named them the cytochromes, or “cellular pigments”, and classified nthese heme proteins, on the basis of the position of their lowest energy nabsorption band in the reduced state, as cytochromes a (605 nm), b n(~565 nm), and c (550 nm). The UV-visible spectroscopic nsignatures of hemes are still used to identify heme type from the reduced nbis-pyridine-ligated state, i.e., the pyridine hemochrome method. Within each nclass, cytochrome a, b, or c, early cytochromes are nnumbered consecutively, e.g. cyt c, cyt c1, and cyt c2, nwith more recent examples designated by their reduced state R-band maximum, ne.g. cyt c559.[2]
Structure and nfunction
The heme ngroup is a highly-conjugated ring system (which allows its electrons nto be very mobile) surrounding a metal ion, which readily interconverts betweethe oxidation states. For many cytochromes, the metal ion present is that of iron, nwhich interconverts between Fe2+ (reduced) and Fe3+ n(oxidised) states (electron-transfer nprocesses) or between Fe2+ (reduced) and Fe3+ (formal, noxidized) states (oxidative processes). Cytochromes are, thus, capable of nperforming oxidation and nreduction. Because the cytochromes (as well as other ncomplexes) are held within membranes in an organized way, the redox nreactions are carried out in the proper sequence for maximum efficiency.
In the process of oxidative phosphorylation, nwhich is the principal energy-generating process undertaken by organisms, other nmembrane-bound and -soluble complexes and cofactors are involved nin the chain of redox reactions, with the additional net effect that protons (H+) nare transported across the mitochondrial inner membrane. The resulting transmembrane proton gradient n([protonmotive force]) is used to generate ATP, which is nthe universal chemical energy currency of life. ATP is consumed to drive cellular nprocesses that require energy (such as synthesis of macromolecules, active ntransport of molecules across the membrane, and assembly of flagella).
Types
Several kinds of cytochrome exist and ncan be distinguished by spectroscopy, exact nstructure of the heme group, inhibitor sensitivity, and reduction potential.
Three types of cytochrome are ndistinguished by their prosthetic groups:
|
Type |
Prosthetic group |
The definition of cytochrome c is not ndefined in terms of the heme group. There is no “cytochrome e,” but nthere is a cytochrome f, nwhich is often considered a type of cytochrome c.
In mitochondria and chloroplasts, these ncytochromes are often combined in electrotransport and related metabolic pathways:
|
Cytochromes |
Combination |
|
a and a3 |
Cytochrome c oxidase (“Complex IV”) with electrons delivered to complex by soluble cytochrome c (hence the name) |
|
b and c1 |
Coenzyme Q – cytochrome c reductase (“Complex III”) |
|
b6 and f |
A completely distinct family of cytochromes is knowas the cytochrome nP450 oxidases, so named for the characteristic Soret peak formed by nabsorbance of light at wavelengths near 450 nm when the heme iron is nreduced (with sodium dithionite) nand complexed to carbon monoxide. nThese enzymes are primarily involved in steroidogenesis and detoxification.
Respiratory chain
The nsystem of mitochondrial enzymes and redox carrier molecules which ferry nreducing equivalents from substrates to oxygen are collectively known as the electrotransport system, or the respiratory chain. This system captures the free energy available nfrom substrate oxidation so that it may later be applied to the synthesis of nATP. Many respiratory chain components were first identified in crude nhomogenates through their spectral properties, which frequently change when a ncarrier is oxidised or reduced. Fractionation of mitochondria in the presence nof mild detergents or chaotropic salts dissected the respiratory chain into nfour large multi-subunit complexes containing the principal respiratory ncarriers, named Complex 1 to Complex 4.
These nsubstantial protein “icebergs” float in the sheet of inner membrane lipids, noften presenting one face to the mitochondrial matrix nand another to the inter – membrane space. nMany of their components have now been isolated in a relatively pure form. nOther membrane bound enzymes such as the energy linked transhydrogenase (ELTH) nare also present which fulfil ancillary roles.
The main components participate in the approximate norder of their redox potentials, and the nbulky complexes are linked by low molecular weight mobile carriers which ferry nthe reducing equivalents from one complex to the next. Except for succinate dehydrogenase n(complex 2) all these complexes pump protons from the matrix space into the ncytosol as they transfer reducing equivalents (either hydrogen atoms or nelectrons) from one carrier to the next. The diagram above shows the flow of nreducing equivalents in purple, and movement of the positively charged protons nin red. Proton pumping is an arduous task which creates substantial pH and nelectrical gradients across the mitochondrial inner membrane. These protons neventually re-enter the matrix space via the F1 ATPase, driving the nsynthesis of ATP as they return.
The number of protons and the number of positive ncharges crossing the inner membrane need not necessarily agree for each nindividual transmembrane protein, although the accounts must balance for the nwhole ensemble. This discrepancy is illustrated on the diagram above, and is nexplained in greater detail below.
Electrons flow through the electrotransport chain to molecular oxygen; during this flow, protons are moved across nthe inner membrane from the matrix to the intermembrane space. This model for nATP synthesis is called the chemiosmotic mechanism, or nMitchell hypothesis. Peter Mitchell, a British biochemist, essentially by nhimself and in the face of contrary opinion, proposed that the mechanism for nATP synthesis involved the coupling between chemical energy (ATP) and osmotic npotential (a higher concentration of protons in the intermembrane space than ithe matrix). The inner membrane of the mitochondrion is tightly packed with ncytochromes and proteins capable of undergoing redox changes. There are four nmajor protein-membrane complexes.
Complex I and Complex II
Complex I and Complex II direct nelectrons to coenzyme Q. Complex I, also called NADH-coenzyme Q reductase, naccepts electrons from NADH. The NADH releases a proton and two electrons. The electrons nflow through a flavoprotein containing FMN and an iron-sulfur protein. First, nthe flavin coenzyme (flavin mononucleotide) and then the iron-sulfur center nundergo cycles of reduction and then oxidation, transferring their electrons to na quinone molecule, coenzyme Q (see Figure ).
Complex I is capable of transferring protons from the nmatrix to the intermembrane space while undergoing these redox cycles. One npossible source of the protons is the release of a proton from NADH as it is noxidized to NAD, although this is not the only explanation. Apparently, nconformational changes in the proteins of Complex I also are involved in the nmechanism of proton translocation during electron transport.
|
|
Complex II, also known as nsuccinate-coenzyme Q reductase, accepts electrons from succinate nformed during the TCA cycle. Electrons flow from succinate to FAD (the flavin-adenine ndinucleotide) coenzyme, through an iron-sulfur protein and a cytochrome b550 nprotein (the number refers to the wavelength where the protein absorbs), and to ncoenzyme Q. No protons are translocated by Complex II. Because translocated nprotons are the source of the energy for ATP synthesis, this means that the noxidation of a molecule of FADH2 inherently leads to less ATP nsynthesized than does the oxidation of a molecule of NADH. This experimental nobservation also fits with the difference in the standard reduction potentials nof the two molecules. The reduction potential of FAD is -0.22 V, as opposed to n-0.32 V for NAD.
Coenzyme Q is capable of accepting either one or ntwo electrons to form either a semiquinone or hydroquinone nform. Coenzyme Q is not bound to a protein; instead it is a mobile electrocarrier and can float within the inner membrane, where it can transfer nelectrons from Complex I and Complex II to Complex III.
Complex III is also known as coenzyme nQ-cytochrome c reductase. It accepts electrons from reduced coenzyme nQ, moves them within the complex through two cytochromes b, an iron-sulfur nprotein, and cytochrome c1. Electron flow through Complex II ntransfers proton(s) through the membrane into the intermembrane space. Again, nthis supplies energy for ATP synthesis. Complex III transfers its electrons to nthe heme group of a small, mobile electron transport protein, cytochrome nc.
Cytochrome c transfers its electrons nto the final electron transport component, Complex IV, or cytochrome noxidase. Cytochrome oxidase transfers electrons through a ncopper-containing protein, cytochrome a, and cytochrome a3, and nfinally to molecular oxygen. The noverall pathway for electron transport is therefore:
|
||
|
|
|
The number n is a fudge nfactor to account for the fact that the exact stoichiometry of proton transfer nisn’t really known. The important point is that more proton transfer occurs nfrom NADH oxidation than from FADH2 oxidation.
Chemiosmotic hypothesis
A theory postulated by the biochemist Peter Mitchell in 1961 to describe nATP synthesis by way of a proton electrochemical coupling is called chemiosmotic nhypothesis.
Accordingly, nhydrogen ions (protons) are pumped from the mitochondrial matrix to the intermembrane nspace via the hydrogen carrier proteins while the electrons are transferred nalong the electron transport chain in the mitochondrial inner membrane. As the nhydrogen ions accumulate in the intermembrane space, an energy-rich protogradient is established. As the proton gradient becomes sufficiently intense nthe hydrogen ions tend to diffuse back to the matrix (where hydrogen ions are nless) via the ATP synthase (a transport protein). As the hydrogen ions diffuse n(through the ATP synthase) energy is released which is then used to drive the nconversion of ADP to ATP (by phosphorylation).
Chemiosmotic nHypothesis in a simple form
In the 1960s, ATP nwas known to be the energy currency of life, but the mechanism by which ATP was ncreated in the mitochondria nwas assumed to be by substrate-level nphosphorylation. Mitchell’s chemiosmotic hypothesis nwas the basis for understanding the actual process of oxidative nphosphorylation. At the time, the biochemical nmechanism of ATP synthesis by oxidative phosphorylation was unknown.
Mitchell realised that the movement nof ions across an electrochemical nmembrane potential could provide the energy needed to nproduce ATP. His hypothesis was derived from information that was well known ithe 1960s. He knew that living cells had a membrane potential; ninterior negative to the environment. The movement of charged ions across a nmembrane is thus affected by the electrical forces (the attraction of positive nto negative charges). Their movement is also affected by thermodynamic nforces, the tendency of substances to diffuse from regions nof higher concentration. He went on to show that ATP synthesis was coupled to nthis electrochemical ngradient.
His hypothesis was confirmed by the ndiscovery of ATP synthase, na membrane-bound protein that uses the potential energy of the electrochemical ngradient to make ATP.
The passage back occurs via a specific protochannel. This passage is coupled to ATP-synthesis, using the potential energy nof the proton gradient for the formation of the third phosphate bond of ATP.
We caow calculate the end result of glucose ndegradation: the oxidation is coupled to a decrease of the free energy; 686 nkcal/mol (= 2881 kJ/mol) are obtained by the complete oxidation of glucose. How much nof this energy can the cell use?
1. nSix mol ATP per mol glucose are ngenerated (substrate chain phosphorylation). This is because all steps after nthe breaking down of fructose-1,6-phosphate have to be counted twice (once for neach of the two resulting C3 molecules), so it is 3 x 2 ATPs. Of nthese six ATPs, two are needed to start glycolysis. That leaves nfour.
2. nDuring the course of glycolysis up to nacetyl-CoA, 2 x 2 NADH + H+ are generated. An additional 3 x 2 NADH n+ H+ and 1 x 2 FADH2 are produced in the citric acid ncycle. One NADH + H+ gives three, one FADH2 two ATPs whefed into the respiratory chain. This sums up to 34 ATPs plus the 4 ATPs of nglycolysis. A total of 38 mol ATP are thus gained by the cell’s degradation of none mol glucose. Since each energy-rich bond of ATP contains 7.3 kcal/mol (= n-30.6 kJ/mol), the 38 ATP equal 277 kcal/mol (ca 1163 kJ/mol). This is 40.6% of nthe theoretically possible gain. The other 59.4 percent are set free as heat. nThis is a very high percentage compared to the gain of technical machines like nsteam or petrol engines that is around or below 20 percent.
The nATP synthase enzymes have been remarkably conserved through evolution. The nbacterial enzymes are essentially the same in structure and function as those nfrom mitochondria of animals, plants and fungi, and the chloroplasts of plants. nThe early ancestory of the enzyme is seen in the fact that the Archaea have aenzyme which is clearly closely related, but has significant differences from nthe Eubacterial branch. The H+-ATP-ase found in vacuoles of the neukaryote cell cytoplasm is similar to the archaeal enzyme, and is thought to nreflect the origin from an archaeal ancestor.
In most nsystems, the ATP synthase sits in the membrane (the “coupling” nmembrane), and catalyses the synthesis of ATP from ADP and phosphate driven by na flux of protons across the membrane down the proton gradient generated by nelectron transfer. The flux goes from the protochemically positive (P) side n(high proton electrochemical potential) to the protochemically negative (N) nside. The reaction catalyzed by ATP synthase is fully reversible, so ATP nhydrolysis generates a proton gradient by a reversal of this flux. In some nbacteria, the main function is to operate in the ATP hydrolysis direction, nusing ATP generated by fermentative metabolism to provide a proton gradient to ndrive substrate accumulation, and maintain ionic balance.
ADP + Pi + nH+P n<=> ATP + nH+N
· nthe nfree energy change (DG) associated nwith synthesis of ATP under cellular conditions (the free nenergy required)
Because the nstructures seen in EM, the subunit composition, and the sequences of the nsubunits appeared to be so similar, it had been assumed that the mechanisms, nand hence the stoichiometries, would be the same. In this context, the evidence nsuggesting that the stoichiometry of H+/ATP (n above) varied ndepending on system was surprising. Values based on measure ATP/2e– nratios, and H+/2e– ratios had suggested thatwas 3 for nmitochondria, and 4 for chloroplasts, but these values were based on the nassumption of integer stoichiometries. Although all the F1F0-type nATP-synthases likely had a common origin, both the assumption that the nstoichiometries are the same, and thatis integer, are called into questioby emerging structural data (see below).
Structure nof the F1 ATP-ase
The structure of the soluble (F1) nportion of the ATP synthase from beef heart mitochondria has been solved by X-ray ncrystallography. The pictures below are from Abrahams, J.P., Leslie, A.G., nLutter, R. and Walker, J.E. n(1994) Structure at 2.8 Å resolution of F1-ATPase from bovine nheart mitochondria
Mechanism of the F1 ATP-ase
The ATP nsynthase operates through a mechanism in which the three active sites undergo a nchange in binding affinity for the reactants of the ATP-ase reaction, ATP, ADP nand phosphate, as originally predicted by Paul Boyer. The change in affinity naccompanies a change in the position of the g-subunit relative to the a, b-ring, nwhich involves a rotation of the one relative to the other. In the direction of nATP synthesis, the rotation is driven by a flux of H+ down the nproton gradient, through a coupling between the g-subunit, and the c-subunit of FO. This nrotation has now been demonstrated experimentally.
Respiratory control
The dependence of oxidative phosphorylation on ADP reveals an important ngeneral feature of this process: Respiration is tightly coupled to the nsynthesis of ATP. Not only is ATP synthesis nabsolutely dependent on continued electron flow from substrates to oxygen, but nelectron flow iormal mitochondria occurs only when ATP is being synthesized nas well. This regulatory phenomenon, called respiratory control, makes nbiological sense, because it ensures that substrates will not be oxidized nwastefully. Instead, their utilization is controlled by the physiological need nfor ATP.
In most naerobic cells the level of ATP exceeds that of ADP by 4- to 10-fold. Respiration ndepends on ADP as a substrate for phosphorylation. When ATP is consumed at high nrates, accumulation of ADP stimulates respiration, with concomitant activatioof ATP resynthesis. Conversely, in a relaxed and well-nourished cell, ATP naccumulates at the expense of ADP, and the depletion of ADP limits the rate of nboth electron transport and its own phosphorylation to ATP. Thus, the nenergy-generating capacity of the cell is closely attuned to its energy ndemands.
Experimentally, respiratory control is demonstrated by following noxygen utilization in isolated mitochondria. In the absence of added substrate or nADP, oxygen uptake, caused by oxidation of endogenous substrates, is slow. nAddition of an oxidizable substrate, such as glutamate or malate, has but a nsmall effect on the respiration rate. If ADP is then added, however, oxygeuptake proceeds at an enhanced rate until all of the added ADP has beeconverted to ATP, and then oxygen uptake returns to the basal rate. This nstimulation of respiration is stoichiometric; that is, addition of twice nas much ADP causes twice the amount of oxygen uptake at the enhanced rate. If nexcess ADP is present instead of oxidizable substrate, the addition of nsubstrate in limiting amounts will stimulate oxygen uptake until the substrate nis exhausted.
Two mechanisms nof the control of respiration and ATP synthesis in mitochondria according to nthe utilization of energy (ATP). The first mechanism of respiratory control is nbased on the proton motive force Δp nacross the inner mitochondrial membrane (grey). Activation of the ATP-synthase n(blue) by ADP, taken up via the ATP/ADP carrier (margenta), decreases Δp which in consequence stimulates the nthree proton pumps of the respiratory chain (complexes I, III and IV). For nsimplicity, only complex IV (cytochrome c noxidase) and its substrate (cytochrome c) nare shown in green and red, respectively. The second mechanism of respiratory ncontrol is based on the intramitochondrial ATP/ADP ratio. High ATP/ADP ratios ninhibit cytochrome c oxidase nactivity allosterically. Uptake of ADP decreases the intramitochondrial ATP/ADP nratio accompanied by exchange of bound ATP by ADP at the matrix domain of nsubunit IV of cytochrome c noxidase, with subsequent stimulation of respiration
Maintenance nof respiratory control depends on the structural integrity of the nmitochondrion. Disruption of the organelle causes electron transport to become nuncoupled from ATP synthesis. Under these conditions, oxygen uptake proceeds at nhigh rates even in the absence of added ADP. ATP synthesis is inhibited, evethough electrons are being passed along the respiratory chain and used to nreduce O2 to water.
Uncoupling of nrespiration from phosphorylation can also be achieved chemically. Chemical nuncouplers such as DNP nor FCCP nact by dissipating the proton gradient. Addition of an uncoupler to nmitochondria stimulates oxygen utilization even in the absence of added ADP. No nphosphorylation occurs under these conditions because there is no ADP to be nphosphorylated.
The phenomenon of respiratory control nis the subject of today’s studio exercise. nAn oxygen electrode may be used to record [O2] in a closed vessel n(diagram p. 804). Electron transfer, e.g., from NADH to O2, is nmonitored by recording the rate of disappearance of O2. At nright is an idealized representation of an oxygen electrode recording while nmitochondria respire in the presence of Pi, along with an electrodonor (e.g., succinate, or a substrate of a reaction that will generate NADH). nThe dependence of respiration rate on availability of ADP, the substrate for the nATP Synthase, is called respiratory control. The respiratory control nratio is the ratio of slopes after and before ADP addition (b/a).
The P/O ratio is the moles of ADP added, ndivided by the moles of O consumed (based on c) while phosphorylating nthe added ADP.
Inhibitors of tissue respiration
Chemiosmosis can be disrupted by a variety of nchemicals. In oxidative phosphorylation, some of these inhibitors are quite ninfamous:
The nelectron transport chain was determined by studying the effects of particular ninhibitors.
Rotenone nis a common insecticide that strongly inhibits the electron transport of ncomplex I.
Rotenone is a natural product obtained from the nroots of several species of plants. Tribes in certain parts of the world beat nthe roots of trees along riverbanks to release rotenone into the water which nparalyzes fish and makes them easy prey.
Amytal is a barbiturate that inhibits the nelectron transport of complex I. Demerol is painkiller that also inhibits ncomplex
Antibiotic. Induces apoptosis, which is not nprevented by the presence of Bcl-2. Inhibits mitochondrial electron transport nspecifically between cytochromes b and c1. All three of these complex I ninhibitors block the oxidation of the Fe-S clusters of complex I.
2-Thenoyltrifluoroacetone nand carboxin specifically block electron transport in Complex II
Antimycin A is an antibiotic that inhibits nelectron transfer in complex III by blocking the transfer of electrons betweeCyt bH and coenzyme Q bound at the QN site. Antibiotic. Induces apoptosis, nwhich is not prevented by the presence of Bcl-2. Inhibits mitochondrial nelectron transport specifically between cytochromes b and c1.
Cyanide, azide and carbon monoxide all inhibit nelectron transport in Complex IV. The all inhibit electron transfer by binding ntightly with the iron coordinated in Cyt a
This complex oxidizes cytochrome c and nalso reduces O2 to H2O. Remember that ncytochromes have heme cofactors — this is important in our discussion of ncyanide and azide. Cytochrome c is a soluble protein and also nis a mobile carrier. Other inhibitors of cytochrome c oxidase will not be ndiscussed here, but are important biologically, such as sulfide, formate, and nnitric oxide.
Azide and cyanide bind to the iron when the irois in the ferric state. Carbon Monoxide binds to the iron when it is in the nferrous state. Cyanide and azide are potent inhibitors at this site which naccounts for there acute toxicity. Carbon monoxide is toxic due to its affinity nfor the heme iron of hemoglobin. Animals carry many molecules of hemoglobin, ntherefore it takes a large quantity of carbon monoxide to die from carbomonoxide poisoning.
Carbon Monoxide
CO competes with oxygen for binding to the reduced form of ncytochrome c oxidase. Once bound to the cytochrome oxidase, oxygen cannot nattach, and electron transport is stopped. CO is a colorless, tasteless, nnon-irritating toxic gas. When inhaled, the toxic gas enters the nbloodstream, depriving the heart and brain of the oxygeecessary to functiocorrectly. Sensing the body’s need for more oxygen, the victim’s heart rate nincreases to pump more blood to the body’s organs. If a person continues to ninhale CO, he or she faces the risk of breathing difficulty, cardiac trauma, nbrain damage, coma and even death.
Animals nhave relatively few molecules of Cyt a3. nConsequently an exposure to a small quantity of azide or cyanide can be nlethal. The toxicity of cyanide is solely from its ability to arrest electrotransport. Cyanide
CN–, like azide, binds to the iron atom of oxidized ncytochrome, preventing binding of oxygen. Again, since cytochrome oxidase nis inhibited, oxygen metabolism is prevented and thus so is energy ngeneration. Cyanide has long been known as a poison, sometimes used iwarfare. As mentioned above, lethal doses cause death in 15 minutes.
How does cyanide, nazide or carbon monoxide cause death?
They all cause similar toxic repercussions (with the exception of CarboMonoxide, which can bind to Hemoglobin causing your body to be unable to bind noxygen properly and causing you to suffocate);
nHowever, these three also cause chemical suffocation. They bind a protein iour electron transport chain (cytochrome c oxidase). The electron transport nchain is our body’s best way to create energy in the form of ATP. Without it, nwe literally have next to no energy. The electron transport chain is the reasowe have pain in our muscles when we work out–if our body doesn’t have oxygein those tissues and can’t make energy, we make lactic acid, which causes our nmuscles to hurt/burn. Anyway, cyanide, azide and carbon monoxide bind this nprotein (cytochrome c oxidase) in the electron transport chain. Doing this ncauses all of the electrons to stop transferring and no energy to be made. This ncauses cell death. The first place to experience cell death from cyanide, azide nand carbon monoxide is your central nervous system–which is made up of your nbrain and spinal cord.
nSo the first thing to die when someone dies of cyanide, azide or carbomonoxide poisoning?
The coupling between electron transport and oxidative phosphorylatiodepends on the impermeability of the inner mitochondrial membrane to nH+translocation. The only way for protons to go from the intermembrane space to nthe matrix is through ATP synthase. Uncouplers uncouple electron transport from noxidative phosphorylation. They collapse the chemiosmotic gradient by ndissipating protons across the inner mitochondrial membrane. All of the uncouplers nshown to the left, collapse the pH gradient by binding a proton on the acidic nside of the membrane, diffusing through the inner mitochondrial membrane and nreleasing the proton on the membranes alkaline side.
Uncouplers of oxidative phosphorylation stimulate nthe rate of electron flow but not
ATP synthesis.
(a)At relatively low levels of an uncoupling nagent, P/O ratios drop somewhat, but the cell can compensate for this by nincreasing the rate of electron flow; ATP levels can be kept relatively normal. nAt high levels of uncoupler, P/O ratios approach zero and the cell cannot nmaintain ATP levels.
(b) As amounts of an uncoupler increase, the P/O nratio decreases and the body struggles to make sufficient ATP by oxidizing more nfuel. The heat produced by this increased rate of oxidation raises the body ntemperature. The P/O ratio is affected as noted in (a).
(c) Increased activity of the respiratory chaiin the presence of an uncoupler requires the degradation of additional energy nstores (glycogen and fat). By oxidizing more fuel in an attempt to produce the nsame amount of ATP, the organism loses weight. If the P/O ratio nears zero, the nlack of ATP will be lethal
2,4-Dinitrophenol, dicumarol and carbonyl ncyanide-p-trifluorocarbonyl-cyanide methoxyphenyl hydrazone (FCCP) all have nhydrophobic character making them soluble in the bilipid membrane. All of these ndecouplers also have dissociable protons allowing them to carry protons from nthe intermembrane space to the matrix which collapses the pH gradient. The potential nenergy of the proton gradient is lost as heat DNP is a chemical nuncoupler of electron transport and oxidative phosphorylation.
DNP npermeabilizes the inner mitochondrial membrane to protons, destroying the nproton gradient and, in doing so, uncouples the electron transport system from nthe oxidative phosphorylation. In this situation, electrons continue to pass nthrough the electron transport system and reduce oxygen to water, but ATP is not nsynthesized in the process. The compound, trifluorocarbonylcyanide nphenylhydrazone (FCCP), is also auncoupler.
The phenolic group of DNP is usually ndissociated at intracellular pH. However, a DNP molecule that approaches nthe inner mitochondrial membrane from the outside becomes protonated (because nthe pH is lower there). Protonation increases the hydrophobicity of DNP, nallowing it to diffuse into the membrane and, by mass action, to pass through. nOnce inside, the higher pH of the matric deprotonates the phenolic hydroxyl nagain. Thus, DNP has the effect of transporting H+ back ninto the matrix, bypassing the F0 proton channel and thereby preventing ATP nsynthesis.
The link between electron transport and ATP synthesis is below. (a) Ithe presence of excess phosphate and substrate and intact mitochondria, oxygeis consumed only when ADP is added. When all of the added ADP has beeconverted into ATP, electron transport stops and oxygen consumption ceases. (b) nThe addition of 2,4-dintrophenol uncouples electron transfer from ATP nsynthesis. The oxygen is completely consumed in the absence of ADP. Endogenous nUncouplers Enable Organisms to Generate Heat
.
The uncoupling of oxidative phosphorylation from nelectron transport generates heat. Hibernating animals and newborne animals n(including human beings) contain brown adipose tissue. The adipose tissue is nbrown due to the high mitochondria content of the tissue. An endogenous proteicalled thermogenin uncouples ATP synthesis from electron transport by opening nup a passive proton channel (UCP-1) through the inner mitochondrial membrane. nThe collapse of the pH gradient generates heat. An uncoupling nprotein (also called thermogenin) is produced in brown adipose tissue nof newborn mammals and hibernating mammals .
This protein of the inner mitochondrial membrane nfunctions as a H+ carrier. The uncoupling proteiblocks development of a H+ electrochemical gradient, thereby nstimulating respiration. The free energy change associated with respiration is ndissipated as heat. This “non-shivering thermogenesis” is costly iterms of respiratory energy unavailable for ATP synthesis, but it provides nvaluable warming of the organism.
Valinomycin ncombines with K ions to form a complex that passes through the inner
Valinomycin is a potent antibiotic which acts as a npotassium (K+) ionophore. Induces K+ nconductivity in cell membranes. Also active in vitro against Mycobacterium nTuberculosis, and as an apoptosis inducer.
Valinomycin is obtained from the cells of several Streptomyces nstrains, among which “S. ntsusimaensis” and S. fulvissimus. nIt is a member of the group of natural neutral ionophores because it does not nhave a residual charge. It consists of enantiomers D- and L-valine (Val), nD-hydroxyvaleric acid and L-lactic acid. Structures are alternately bound via namide and ester bridges. Valinomycin is highly selective for potassium ions nover sodium ions within the cell membrane. It functions as a potassium-specific ntransporter and facilitates the movement of potassium ions through lipid nmembranes “down” an electrochemical potential gradient. The stability nconstant K for the potassium-valinomycin complex is 106 and for the nsodium-valinomycin complex only 10. This difference is important for maintaining nthe selectivity of valinomycin for the transport of potassium ions (and not nsodium ions) in biological systems.
Oligomycin is a natural nantibiotic isolated from Streptomyces diastatochromogenes which ninhibits mitochondrial H+-ATP synthase. It is primarily found to act nas an inhibitor of mitochondrial respiration and swelling. This nantibiotic is widely used as an inhibitor of oxidative phosphorylation.
nOligomycin inhibits the H+– ATP-synthase by binding to the Oligomycisensitivity-conferring protein (OSCP) at the F(o) subunits 6 nand 9 which are found in the stalk of the F1F0-ATPase ncomplex. This binding blocks the proton conductance and inhibits the synthesis nof mitochondrial ATP.3-4 Because of its activity, it can also be nused to reduce the number of parameters (such as ER Ca2+ nrelease, exocytotoxicity and apopotosis) which are affected by mitochondrial ndepolarization.
n
nOligomycin, at high concentrations may also inhibit the plasma membrane nNa+-K+-ATPase. Interaction of Oligomycin with the Na+ nocclusion site on the extracellular side of Na/K-ATPase, delays Na+ nrelease to the extracellular side without inducing a conformational change. nAlthough Oligomycin stimulated Na+ binding to Na+/K+-ATPase, nit inhibited Na+/Na+ exchange, and did not affect neither Na+-dependent AD/-ATP exchange or K+-dependent nphosphatase activity.6-16
Proton Motive Force Drives Transport The primary purpose of the protogradient is togenerate ATP by oxidative phosphorylation. The potential energy nof the gradient can also be used for active transport. The inner mitochondrial nmembrane is impermeable to charged molecules.
ATP-ADP Translocase There are two specific systems to transport ADP and nPi into the mitochondrial matrix. A specific transport protein ATP-ADP ntranslocase enables ATP and ADP to transverse the inner mitochondrial membrane. nThe transport of ADP in and ATP out are coupled. ADP only enters the matrix if nATP exits or vice versa. ATP-ADP translocase has a single nucleotide binding nsite which binds ADP and ATP with equal affinity. Due to the negative nelectrostatic charge of the matrix, ATP-is bound on the N phase of the membrane nbecause it has greater negative charge than ADP.
Pentachlorophenol n(PCP) acts in a similar way to DNP. It was widely used nas a biocide, especially in pallet board manufacture as a fungicide, but is now nbanned by the Biocidal Products Directive, because of its extreme toxicity and nenvironmental persistence.
nDNP
nPCP
Ways of nenergy usage in the organism.
o n1. From the smallest, single-celled organism nto the biggest and most complex mammals–including people–all living things nrequire energy for life. It’s easy enough to understand that we and other nanimals eat. Things get a little more puzzling when we think about fungi, which nabsorb their food as organic molecules, from the surrounding environment. Where ndo those molecules come from? Furthermore, where does the food come from that nwe humans convert to energy? At the most basic level, all energy traces back to nplants. Plants are the basis of all the world’s food systems, and their unique nability to make organic materials from sunlight–called photosynthesis–is what nsustains nearly every other life form on the planet.
o nThe powerhouse of energy productioin all plants is called a chloroplast. More than a million of these handy ndevices occur in every quarter-inch of a leaf. They contain the pigment called nchlorophyll that makes most leaves green–and drives photosynthesis. The nreaction isn’t all that complicated, as far as chemical reactions go. The nchloroplasts take in carbon dioxide, sunlight and water. They release oxygeand a bit less water than they took in. The conversion of carbon dioxide to noxygen is one life-sustaining function that plants perform for Earth and all of nits life. But plants do something equally as important when they keep a third nproduct behind: glucose, the sugar that sustains the plants—and anything, iturn, that eats the plants.
o nIn cellular respiration, glucose is nbroken down by the removal of its hydrogen atoms. That process releases energy nin the form of electrons, negatively charged particles that fuel all of a ncell’s other work in later reactions. So, plants make the glucose and neverything down the line—from plant-eaters to the carnivores that eat nthem—break the glucose down again, and use its energy. That’s the simple nstory. Of course, life is rarely so simple, and there are exceptions to every nrule. Every so often, a new discovery comes along about living things that use na non-living substance other than sunlight to make energy–like ammonia, or neven sulfur. These less-common organisms can harness electrons from chemical nsources instead of the sun. More amazing life forms have the potential to be ndiscovered at any time, anywhere on our planet—or beyond.
Cytochrome P450
The active site of cytochrome P450 ncontains a heme niron center. The iron is tethered to the P450 protein via a thiolate nligand derived from a cysteine nresidue. This cysteine and several flanking residues are highly conserved iknown CYPs and have the formal PROSITE nsignature consensus pattern [FW] – [SGNH] – x – [GD] – {F} – [RKHPT] – {P} – C n- [LIVMFAP] – [GAD].[7] nBecause of the vast variety of reactions catalyzed by CYPs, the activities and nproperties of the many CYPs differ in many aspects.

