БІОЕНЕРГЕТИЧНІ ПРОЦЕСИ: БІОЛОГІЧНЕ ОКИСНЕННЯ,
ТКАНИННЕ ДИХАННЯ. МОЛЕКУЛЯРНА ОРГАНІЗАЦІЯ МІТОХОНДРІАЛЬНОГО ДИХАЛЬНОГО ЛАНЦЮГА.
ХЕМІОСМОТИЧНА ТЕОРІЯ ОКИСНОГО ФОСФОРИЛЮВАННЯ. ІНГІБІТОРИ І РОЗ’ЄДНУВАЧІ
ОКИСНОГО ФОСФОРИЛЮВАННЯ
Вузлові питання біоенергетики
Розділ біохімії, що
займається питаннями перетворення і використання енергії у живих системах,
називається біоенергетикою. Біоенергетика грунтується на процесах
окиснення органічних речовин.
Початки вивчення енергетичних процесів
у живих організмах були закладені французьким ученим Лавуазьє ще в XVIII ст.
Він уперше звернув увагу на подібність згорання органічних речовин поза
організмом з процесом
дихання в тварин. Оскільки при згоранні речовин утворюються СО2 і Н2О,
а під час дихання виділяються такі ж речовини і певна кількість тепла, то
Лавуазьє цілком логічно вважав, що дихання організму – процес з'єднання кисню
повітря з вуглецем і воднем органічних речовин всередині тіла. Експерименти
Лавуазьє, а також його висновки послужили основою для ствердження, що живі
організми підпорядковуються першому принципу термодинаміки. Це означає, що
спалювання цукру поза організмом (in vitro) і окиснення його в організмі (in
vivo) можна зобразити у вигляді хімічної реакції:
С6Н12O6 + 6О2
® 6СО2
+ 6 Н2О + тепло
Всі хімічні реакції в живих
клітинах підпорядковуються законам термодінамики.
Перший закон: енергія не створюється з нічого
та не зникає, вона лише перетворюється в еквівалентних кількостях з одного виду
в інший, тобто внутрішня енергія системи залишається постійною. Самовільно
хімічні процеси можуть проходити лише в одному напрямку – досягнення
рівноваги, тобто максимальної ентропії.
Ентропія – це міра хаосу, неупорядкованості.
Другий закон: ентропія при
самовільних процесах зростає. Самовільно неупорядкована система ніколи
не
перетвориться в
упорядковану. Для цього перетворення треба
витратити додаткову енергію. Напрямок перебігу реакцій можна визначити на
практиці за допомогою вільної енергії системи, котру можна виміряти. Енергетичний стан
будь-якої системи виражається цією формулою:
D G = D Н - T D S
D G - це та частина енергії системи, котру можна використовувати для виконання роботи; Т D
S - зв’язана енергія, котра не може бути
використана на роботу; DН - ентальпія – внутрішня енергія системи (може бути вимірена експериментально). - якщо DG < 0, то реакція екзергонічна,
тобто перебігає самовільно з вивільненням енергії (ці
реакції постачають клітинам енергію); - якщо DG > 0, то реакція ендергонічна,
тобто відбувається з використанням зовнішньої енергії; - якщо DG = 0, - це стан рівноваги.
Екзергонічні і ендергонічні
процеси
Життєво необхідні процеси –
реакції синтезу, скорочення м’язів, проведення нервових імпульсів, активний транспорт – отримують енергію шляхом хімічного
поєднання з окисними реакціями, тобто існує поєднання екзергонічних реакцій з
ендергонічними.
До екзергонічних реакцій
належать катаболічні
реакції - реакції
розщеплення або окиснення “паливних”
молекул (енерговмісних нутрієнтів), котрі надходять в організм у складі
харчових продуктів.
До ендергонічних реакцій належать анаболічні реакції - реакції
синтезу складних біоорганічних сполук – клітинних макромолекул.
Сукупність усіх біохімічних перетворень хімічних речовин у
живому організмі - це метаболізм. Для поєднання цих процесів
необхідний посередник (рис. 1). У
процесі екзергонічних реакцій синтезуються сполуки з високим енергетичним потенціалом ~ Е, котрі вже далі
використовуються в ендергонічних реакціях. Таким шляхом відбувається передача
хімічної енергії від екзергонічного до ендергонічного процесу. Сполуки з
високим енергетичним потенціалом, макроергічні сполуки (~Е), – це біомолекули з
високою стандартною вільною енергією
перенесення кінцевої фосфатної групи.

Рис. 1. Поєднання ендергонічних процесів з
екзергонічними
В біології міцно
закріпилось уявлення, що дихання в організмі (окиснення) за своєю суттю є
процесом “горіння", яке відбувається дуже повільно. Отже, поживні
речовини, що потрапили в організм з їжею, є “паливом", яке згорає в
організмі шляхом приєднання кисню повітря. Разом з тим, було звернуто увагу на той факт, що повільне “горіння"
органічних речовин в організмі істотно відрізняється від такого, що проходить
поза організмом: по-перше, воно відбувається при низькій температурі, по-друге,
– при відсутності полум'я і, по-третє, – за наявності води, вміст якої в
тканинах досить високий.
Для пояснення перебігу окисних процесів
в організмі (повільне горіння) були запропоновані гіпотези активації кисню в
клітинах. Першу спробу пояснити механізм активації кисню в організмі здійснив
німецький хімік Христіан Фредерік Шенбайн (1860 р.), котрий спершу відкрив
активний кисень – озон, що має здатність окиснювати речовини і при
звичайній температурі. На цій підставі Шенбайн припустив, що в організмі під
впливом легкоокиснювальних речовин відбувається розщеплення молекули кисню на
два атоми озону, які легко окиснюють інші речовини. Одначе гіпотеза Шенбайна не
була підтверджена експериментально. Значення праць Шенбайна тільки в тому, що
вони стали початком наступних теорій активації кисню в організмі.

Христіан Фрідріх Шeнбaйн (1799—1868)
У 1897 р. була
обгрунтована перша гіпотеза біологічного окиснення, названа гіпотезою
перекисного (пероксидного) окиснення. Її запропонували одночасно російсько-український учений О.М. Бах та
німецький Енглер. Ці автори вважали, що молекула кисню в тілі живого активується
легкоокиснювальними речовинами, але за іншими механізмами, ніж це передбачав
Шенбайн.
. 
Олексій Миколайович Бах (1857 – 1946)
Припускалось, що в
процесі активації кисню відбувається розрив не обох зв'язків, а тільки одного,
і в подальшому окиснення відбувається шляхом приєднання до речовини всієї
молекули кисню з утворенням молекули пероксиду:
![]()
![]()
Кисень у цій реакції приєднується до
органічної речовини (позначеної буквою А), що має здатність до автоокиснення –
оксигенази. Далі утворений пероксид віддає активований кисень субстратові
окиснення за допомогою ферменту пероксидази. Таким чином, у процесі окиснення, за
пероксидною гіпотезою, необхідно два ферменти – оксигеназа, яка активує кисень
з утворенням пероксиду, та пероксидаза, яка окиснює речовину за допомогою
пероксиду. Але пізніше було показано, що цей механізм є не головним, а тільки
окремим випадком біологічного окиснення, і не має відношення до
енергозабезпечення тканин. Такий тип окиснення органічних речовин має місце в
ендоплазматичному ретикулумі печінки, і його призначення – знешкоджувати
шляхом окиснення токсичні речовини.
Ідею про активацію кисню в процесі
біологічного окиснення продовжив і значно розвинув знаменитий німецький учений
О. Варбург. Він розробив вчення, згідно з яким активація кисню є ключовою
ланкою в біологічному окисненні, результатом якого є з'єднання кисню і водню з
утворенням води. Варбургу належить також відкриття гемвмісного ферменту –
цитохромоксидази, що активує кисень у процесі біологічного окиснення, він же
вперше сконструював апарат для вивчення тканинного дихання, названий його
іменем.
Треба зазначити,
що одночасно з відкриттям цитохромоксидази були відкриті ферменти
дегідрогенази, які здатні відщеплювати від субстратів атоми водню і цим самим
окиснювати їх.

Отто Варбург (1883 – 1970)
Була запропонована нова гіпотеза
біологічного окиснення, згідно з якою визначальна роль відводилась процесам
дегідрування субстратів (відщеплення атомів водню). Цю ідею відстоювали
Паладін, Віланд, Тунберг та ін. Тільки з відкриттям у 1933 р. Кейліним
цитохромів, які виявились проміжними переносниками електронів від водню до
кисню, були узгоджені оксидазна концепція Варбурга з дегідрогеназною Паладіна,
Віланда.

Володимир Іванович Паладін (1859 –
1922)
Це означало, що як
активація атомів водню в субстраті за допомогою дегідрогеназ, так і активація
кисню за участю цитохромоксидази є необхідними складовими ланками біологічного
окиснення, яке відбувається в тканинах організму. Таким чином, теорія біологічного
окиснення, вперше запропонована у XVIII ст. Лавуазьє як процес повільного
горіння, що призводить до утворення СО2, Н2О і енергії,
була завершена тільки в XX ст. В опрацюванні її брали участь також Кейлін,
Кребс, Мітчел, Ленінджер, Чанс, Рекер, Беліцер, Енгельгардт та ін. Цими ж
авторами було доведено, що окиснення субстратів до СО2 і Н2О
з виділенням енергії відбувається в різних тканинах організму, тому біологічне
окиснення називають ще тканинним або клітинним диханням. Основним місцем у
клітині, де завершується окиснення субстратів та виділяється енергія, є
мітохондрії, тому їх назвали енергетичними станціями клітини. В мітохондріях
відбувається не тільки окиснення субстратів шляхом відщеплення атомів водню і
перенесення останніх на кисень, але й одночасне перетворення та запасання
енергії у хімічній формі в так званих макроергічних зв'язках аденозинтрифосфату
(АТФ).
|
Біологічне
окиснення – це сума всіх окисно-відновних процесів, включаючи анаеробні, що
відбуваються в клітинах організму (цитоплазмі, мітохондріях,
ендоплазматичному ретикулумі). Основними субстратами
біологічного окиснення є вуглеводи та ліпіди, саме їх катаболізм дає
найбільшу кількість хімічної енергії, що акумулюється у високоенергетичних
зв’язках макроергів. |
Функції біологічного окиснення:
1. Забезпечення клітин енергією;
2. Забезпечення клітин пластичним
матеріалом для відтворення структур організму;
3. Знешкодження токсичних речовин.
|
Тканинне дихання – це окиснення
речовин з допомогою кисню, що відбувається на внутрішній мембрані
мітохондрій, супроводжується утворенням Н2О і АТФ. |
|
Дихальний ланцюг або дихальний ансамбль – це система ферментів і
коферментів, з допомогою яких відбувається транспорт електронів і протонів
водню від субстрату до кисню з утворенням Н2О і АТФ. Його складові вбудовані у внутрішню
мембрану мітохондрій: від 5 до 20 тисяч ансамблів в одній мітохондрії. |
|
Енергетичний
обмін – це перетворення енергії харчових речовин в інші форми хімічної
енергії (АТФ, НАДН, ФАДН2) і використання цих форм енергії для виконання
роботи. |
Біологічне окиснення каталізується
ферментами і може відбуватися такими
шляхами:
1) приєднання кисню до субстрату
окиснення;
2) відщеплення водню від субстрату
(дегідрування);
3) відщеплення електронів.
Біоенергетика включає 3 вузлові
питання:
1) джерела енергії;
2) способи перетворення і нагромадження
енергії;
3) шляхи використання енергії.
Типи реакцій біологічного
окиснення: дегідрогеназні, оксигеназні,
оксидазні, пероксидазні, вільнорадикальні,
їх суть і біологічне призначення.
Існують різні типи
реакцій окиснення органічних субстратів, в котрих використовується кисень :
1.
Дегідрогеназні реакції. У
результаті цих реакцій відбувається перенесення водню з субстрату (S) на
акцептор (А):
SH2 + A → S + AH2.
Ферменти, що каталізують реакції цього класу, мають
назву дегідрогенази.
Залежно від типу акцептора водню реакції
дегідрування поділяють на два підкласи:
- реакції, які каталізують анаеробні дегідрогенази,
акцептором (А) є сполука, яка відмінна від кисню (НАД, НАДФ, ФАД, ФМН,
убіхінон, глутатіон, ліпоєва кислота тощо);
- реакції,
які каталізують аеробні дегідрогенази (оксидази), акцептором (А) в цих реакціях
є кисень (О2):
SH2 + 1/2O2 → S + H2O
(цитохромоксидаза)
Оксидазні
реакції (80-90% споживання кисню). Продукти – окиснений субстрат, вода та енергія. Реалізується у мітохондріях у дихальному
ланцюзі.
2.
Оксигеназні реакції (8-12% споживання кисню). Реакції
цього класу каталізують ферменти оксигенази, які приєднують до субстрату
окиснення один або два атоми кисню. Тому залежно від кількості атомів кисню, що
приєднуються, ферменти поділяють на:
-
монооксигенази (відповідно реакції –
монооксигеназні):
SH + 1/2О2 →
S-ОН
До монооксигеназ належить цитохром Р450,
що каталізує гідроксилювання ендогенних та екзогенних субстратів;
- диокисгенази (відповідно реакції – диоксигеназні):
S + О2
→ SО2.
- за допомогою монооксигеназ:
один атом кисню включається
у
субстрат, другий – у молекулу води; за допомогою диоксигеназ: обидва атоми кисню включаються у субстрат.
3. Пероксидазні
реакції: це шлях відновлення кисню до пероксиду водню за допомогою пероксидутворюючих ферментів –
флавопротеїнів.
SH2 + О2 → S + H2О2
(ксантиноксидаза, аміноксидаза,
глюкозоксидаза)
За цим шляхом
окиснюються біогенні аміни, амінокислоти. Перекис водню – це токсична речовина,
але у фагоцитуючих клітинах, наприклад, лейкоцитах, її утворення необхідне для
знешкодження патогенних бактерій.
4. Вільнорадикальні реакції відбувається під дією активних форм кисню (АФК), що утворюються шляхом одноелектронного відновлення кисню.
До АФК належать:
- супероксид-аніон;
- перекис водню;
- гідроксильний радикал;
- синглетний кисень.
Макроергічний
зв’язок, макроергічні сполуки
АТФ
відноситься до сполук, що містять макроергічні зв'язки, тобто багаті енергією.
(рис. 2). Зазначимо відмінність поняття
“багатий енергією зв'язок" у біохімії від поняття “енергія зв'язку" у
хімії. Під останнім поняттям у хімії розуміють енергію, необхідну для розриву
зв'язку між двома атомами в молекулі (для розриву макроергічних зв'язків також
потрібно затратити енергію). Розглядаючи в біохімії високоенергетичні та
низькоенергетичні зв'язки і сполуки, енергію зв'язку визначають як вільну
енергію, що виділяється при
гідролітичному розпаді даної сполуки.

Рис. 2.
Хімічна формула молекули АТФ
Енергія
може бути акумульована в різних субстратах окиснення, проте лише енергія у
формі АТФ може бути використана клітинами організму. Саме тому АТФ називають
«універсальним джерелом енергії». Крім
АТФ, існують й інші макроерги. Це метаболіти вуглеводного, ліпідного та
амінокислотного обмінів, а також фосфагени (креатинфосфат), котрі виступають як
резервуари макроергічних зв’язків.
У клітинах містяться різні біофосфати з різним енергетичним потенціалом.
Залежно від кількості енергії, яка акумульована в макроергічних зв’язках цих
сполук, усі біофосфати умовно можна поділити на три групи.
1. Надвисокоенергетичні
фосфати – це фосфати, енергія гідролізу фосфодиефірних зв’язків яких
∆Go> - 30 кДж/моль (або -7,3
ккал/моль).
До цієї
групи належать:
-
фосфоенолпіруват (∆Go= - 61,9
кДж/моль),
- 1,3
–дифосфогліцерат (∆Go= - 54,5
кДж/моль),
-
карбамоїлфосфат (∆Go = - 51, 4 кДж/моль),
-
креатинфосфат (∆Go= - 43,1
кДж/моль) та ін.
Реалізація енергії надвисокоенергетичних фосфатів здійснюється виключно
через синтез АТФ. Наприклад, енергія, що акумульована в креатинфосфаті,
спочатку передається на АТФ, і лише потім використовується:
Креатинфосфат + АДФ ↔ креатин + АТФ
(креатинфосфокіназна реакція)
Далі енергія АТФ буде використана відповідно до потреб організму.
Незважаючи на те, що ∆Go макроергічних зв’язків
АТФ менша, ніж органічних фосфатів цієї групи, лише енергія гідролізу АТФ
безпосередньо може бути використана в ендергонічних реакціях клітини.
2. Високоенергетичні фосфати – це
фосфати, енергія гідролізу фосфодіефірних зв’язків яких приблизно дорівнює -
30кДж/моль (∆Go≈ - 30
кДж/моль). До цієї групи належать нуклеозидтрифосфати, такі як АТФ, ГТФ та ін.
3. Низькоенергетичні фосфати – це
органічні фосфати з ∆Go< - 30
кДж/моль. Енергія низькоенергетичнихфосфатів не може бути використана в ендергонічних процесах.
До цієї групи сполук належать:
- АМФ (∆Go= - 9,6
кДж/моль),
- АДФ (∆Go= - 27,6
кДж/моль),
- гліцерофосфат (∆Go= - 9,2
кДж/моль),
- глюкозо-6-фосфат (∆Go= - 13,8
кДж/моль)
У реакції АТФ + Н2О
® АДФ +
Фн зміна вільної енергії
дорівнює 34,5 кДж/моль. Зміни для інших реакцій гідролізу АТФ, АДФ і АМФ становлять:

Зв'язок вважається високоенергетичним,
якщо при гідролізі його звільняється більше 21 кДж (за іншими джерелами – 30
кДж/моль). Таким
чином, у молекулі АТФ є два макроергічних зв'язки, які характеризуються
величиною вільної енергії 28-37 кДж/моль. Макроергічний зв'язок містить і АДФ,
але при його гідролізі до АМФ і Фн
енергія вилучається у вигляді тепла.
Виходячи з того, що молекула АТФ містить два високоенергетичні
зв’язки, виділення енергії може
відбуватися у вигляді двох реакцій:
АТФ → АДФ + Фн (∆Go= - 30
кДж/моль);
АТФ → АМФ + ФФн (∆Go= - 30
кДж/моль).
Зазначимо, що в різних джерелах
наводяться різні значення ∆G гідролізу АТФ. Це пояснюється тим, що реальне значення вільної енергії
гідролізу АТФ (та інших сполук) залежить від концентрації АТФ, АДФ, АМФ, Фн,
іонів Мg2+, величин рН і температури. При стандартних умовах (рН
7,0; температура 25 °С; концентрація АТФ, АДФ і Фн рівні
Чому при гідролізі АТФ та інших
макроергічних сполук виділяється значно більше енергії, ніж, наприклад, при
гідролізі глюкозофосфатів? Це зумовлено властивостями всієї структури
макроергічної сполуки, а не тільки зв'язком, при гідролітичному розриві якого
виділяється вільна енергія. Існує ряд факторів, які вносять свій вклад у процес
звільнення великої кількості вільної енергії при гідролізі макроергічних
зв'язків:
1) електростатичне відштовхування. При
рН 7,0 у молекулі АТФ всі чотири групи ОН зв'язані з фосфором і несуть
негативні заряди (АТФ4‑). Відштовхування однойменних зарядів
призводить до електростатичної напруги у всій молекулі, особливо зв'язку Р-О-Р.
Коли зв'язок розривається, електростатична напруга знімається за рахунок
просторового роз'єднання негативно заряджених продуктів гідролізу АДФ3-
і НРО42-;
2) фактор конкурентного резонансу. Два
продукти гідролізу, АДФ3- і НРО42-, є
резонансними гібридами, тобто такими структурними формами, що мають підвищену
стійкість, бо частина їх електронів перебуває в конфігураціях з нижчими
енергетичними рівнями, порівняно з АТФ;
3) виділення протонів. Сумарне рівняння
гідролізу АТФ має такий вигляд:
АТФ 4- + Н2О
® АДФ3-
+ НРО42- + Н+
Зв'язування цього протона з буферним
середовищем (рН 7,0) зсуває реакцію вправо, що вносить вклад у зміну вільної енергії.
При гідролізі низькоенергетичних фосфатів ці фактори незначні або взагалі не
спостерігаються.
Інтенсивність оновлення АТФ в організмі
Яка кількість АТФ синтезується і
розпадається за добу? Доросла здорова людина масою
В організмі людини міститься всього
близько
Шляхи синтезу
АТФ в клітинах: субстратне та окисне
фосфорилювання,
їх відмінності.
Синтез АТФ із АДФ
і фосфату (фосфорилювання АДФ)
відбувається в організмі за двома шляхами, котрі відрізняються один від іншого джерелом енергії для утворення макроергічного зв’язку :
Окисне
фосфорилювання (головний шлях синтезу): здійснюється за рахунок енергії
окиснення електронів і протонів водню, відщеплених від різноманітних сполук
(метаболітів або субстратів окиснення),
який відбувається за участю дихального ланцюга мітохондрій за рахунок
електрохімічного потенціалу внутрішньої мембрани. Реакції цього процесу
відбуваються виключно в аеробних умовах. Основна кількість АТФ в організмі утворюється саме в реакціях окисного
фосфорилювання.
Субстратне фосфорилювання - процес синтезу АТФ, який відбувається як результат
окиснення субстратів без участі дихального ланцюга мітохондрій. У цьому разі
перетворення субстрату в продукт супроводжується фосфорилюванням АДФ з
утворенням АТФ. В організмі є три реакції субстратного фосфорилювання (дві – в
гліколізі, одна – в ЦТК). Кожна з цих реакцій супроводжується утворенням лише
одної молекули АТФ. Цей процес можливий як в аеробних, так і анаеробних умовах
і відбувається в цитоплазмі і матриксі мітохондрій.
Найбільш потужними процесами продукування енергії в
клітинах є аеробні. Використання клітинами кисню необхідне для окиснення
субстратів, кінцевими продуктами якого є Н2О, СО2, АТФ.
Крім того, виділяється аміак, який у подальшому зазнає знешкодження. Таким чином,
клітини, тканини «дихають» і виробляють енергію, яка необхідна для
життєдіяльності.
http://www.youtube.com/watch?v=XmrwRAytaMU&feature=related
Хімічна енергія
органічних речовин їжі (вуглеводів, жирів, білків) перетворюється в енергію
макроергічних зв'язків АТФ у процесі тканинного (клітинного) дихання. Суть його зводиться до окиснювального
розщеплення молекул органічних речовин, що супроводжується споживанням О2
з виділенням Н2О і СО2 і звільненням енергії. На першому
і другому етапах катаболізму вуглеводів, жирів і білків звільняється невелика
кількість енергії, відповідно близько 1 і 10 %, причому на першому етапі вона
розсіюється у вигляді тепла, а на другому частково використовується для синтезу
АТФ (у процесі субстратного фосфорилювання під час гліколізу). Основна маса
енергії звільняється на третьому етапі окиснення поживних речовин, що
реалізується у мітохондріях і включає реакції циклу лимонної кислоти (циклу
Кребса) та ланцюг перенесення електронів і протонів на кисень (дихальний
ланцюг). У циклі лимонної кислоти вуглецеві атоми субстратів окиснюються і
звільняються у вигляді СО2.
Окиснення вуглецю відбувається за
рахунок кисню самих субстратів і кисню води. Атоми водню (електрони і протони)
відриваються від субстратів і переносяться компонентами дихального ланцюга на
молекулярний кисень, відновлюючи його до води. Вільна енергія, що звільняється
в дихальному ланцюгу під час електронного перенесення, використовується для
синтезу АТФ із АДФ і Фн
шляхом окиснювального фосфорилювання.
http://www.youtube.com/watch?v=9p6bDf3lu6s&feature=related
Отже, процес біологічного окиснення субстратів – проміжних продуктів
метаболізму вуглеводів, жирів і амінокислот – відбувається не за рахунок
приєднання атомів кисню до молекули речовини, а шляхом відщеплення атомів водню
(дегідрування) і багатоетапного
перенесення електронів і протонів на молекулярний кисень.
Як сказано вище, в
організмі мають місце і реакції окиснення субстратів, коли атоми кисню
включаються у молекулу субстрату або коли субстрат реагує безпосередньо з О2.
Але при таких реакціях неможливий
синтез АТФ. Енергія окиснення при цьому використовується для знешкодження
токсичних або біосинтезу нових речовин.
Ферменти
біологічного окиснення в мітохондріях: піридинзалежні, флавінзалежні
дегідрогенази, цитохроми, їх будова і функції.
Ферменти, які каталізують окиснення субстратів шляхом дегідрування,
називаються дегідрогеназами. Розрізняють дві групи дегідрогеназ:
нікотинаміддинуклеотидні (піридинові) і флавінові. Окиснення більшості
субстратів каталізують ферменти першої групи.
Піридинзалежні дегідрогенази
1. Піридинзалежні дегідрогенази –
дегідрогенази, які містять нікотинамідні коферменти (НАД+ або НАДФ+)
(нікотинамід, що входить до складу цих коферментів, належить до похідного
піридину). Піридинзалежні дегідрогенази – це аеробні і анаеробні
дегідрогенази, що каталізують різноманітні реакції в нашому організмі.
Схематично реакції, що каталізують ці ферменти, мають такий вигляд:
SH2 + НАД+→S + НАДН
+ Н+; та SH2 + НАДФ+→S + НАДФН
+ Н+.
НАД-залежні дегідрогенази – це ферменти
окисно-відновних реакцій процесів катаболізму, таких як гліколіз, цикл Кребса,
β-окиснення жирних кислот, дихальний ланцюг мітохондрій тощо.
НАДФ-залежні дегідрогенази – це ферменти
процесів відновного синтезу: синтезу жирних кислот, холестеролу, стероїдних
гормонів, жовчних кислот тощо.
НАД і НАДФ – це
коферментні форми вітаміну РР-нікотинаміду. Будова НАД і НАДФ показана на (рис 3).


Рис. 3.
Будова НАД і НАДФ
Два нуклеотиди
з'єднані залишками фосфорної кислоти, на відміну від міжнуклеотидного зв'язку в
нуклеїнових кислотах (3I,5I-фосфодіефірного).
Азотною основою в одному нуклеотиді є
аденін, а в другому – нікотинамід. НАДФ містить додаткову фосфатну групу при
С2 рибозного залишку, зв'язаного з аденіном.
Нікотинаміддинуклеотидні дегідрогенази
каталізують зворотні реакції, які в загальному вигляді можуть бути записані
так:
![]()
НАД+ і НАДФ+ – це
окиснені форми коферментів, в яких позитивний заряд несе атом азоту
піридинового кільця нікотинаміду. НАДН і НАДФН – відновлені форми. Піридинове
кільце нікотинаміду є тією частиною НАД (Ф), яка бере участь у переносі водню:

Як видно зі схеми, субстрат втрачає два
атоми водню, тобто 2 протони, 2 електрони, але на кофермент переносяться 2
електрони і 1 протон, а другий протон переходить у середовище. Таким чином,
кофермент отримує два відновні еквіваленти: один із них у формі атома водню
приєднується до четвертого вуглецевого атома нікотинамідного кільця, а другий у
формі електрона передається азоту цього кільця. В результаті відновлена форма
коферменту втрачає позитивний заряд. Зазначимо, що, хоч у дійсності на
кофермент переносяться гідрид-іон Н- (протон і 2 електрони), НАД+
(НАДФ+) називають частіше акцептором електронів, а іноді й
акцептором водню.
Нікотинаміддинуклеотиди знаходяться у
дисоційованій формі і зв'язуються з апоферментом тільки в ході реакції.
Відновлені коферменти, відділившись від дегідрогеназ, знову окиснюються шляхом
перенесення електронів до акцептора, зв'язаного з іншим ферментом. При цьому
НАДН і НАДФН по-різному використовуються у метаболізмі.
Більша частина клітинних дегідрогеназ
переносить водневі атоми від субстратів на НАД+, а відновлений НАДН
передає електрони на дихальний ланцюг. Енергія, що звільняється при передачі
електронів у дихальному ланцюгу, запасається у формі АТФ. Компоненти дихального
ланцюга вмонтовані у внутрішній мембрані мітохондрій, і більшість НАД-залежних
дегідрогеназ локалізовані у матриксі мітохондрій. НАДФ-залежні дегідрогенази
знаходяться у цитоплазмі і мітохондріях, але НАДФН не віддає електрони на
дихальний ланцюг, а використовується як відновник у процесах синтезу багатьох
сполук, зокрема жирних кислот, стероїдів.
Флавінзалежні дегідрогенази
Флавінзалежні дегідрогенази –
дегідрогенази, які містять похідні вітаміну В2 – ФАД, ФМН. У
більшості дегідрогеназ ці коферменти
мають високу спорідненість до білкової частини і не відщеплюються від неї.
Схематично реакції,
що каталізують ці ферменти, мають такий вигляд:
SH2 + ФАД→S + ФАДН2;
та SH2 + ФМН→S + ФМН-Н2.
Флавінзалежні дегідрогенази можуть бути як анаеробними, так і аеробними
дегідрогеназами.
До флавінзалежних дегідрогеназ належать: сукцинатдегідрогеназа (фермент
циклу Кребса), НАДН-дегідрогеназа дихального ланцюга мітохондрій,
гліцерол-3-фосфатдегідрогеназа (фермент окиснення гліцеролу),
ацил-КоА-дегідрогеназа (фермент β-окиснення жирних кислот), оксидази L- та D-амінокислот,
ксантиноксидаза (фермент катаболізму пуринових нуклеотидів), глюкозоксидаза
(рослинний фермент, який використовується для визначення концентрації глюкози в
крові).
За структурою ця група ферментів є
складними білками, флавопротеїнами.

R – залишок рибітолфосфату у ФМН, у ФАД він зв’язується з аденіловою кислотою.
На противагу нікотинамідним
коферментам, ФМН і ФАД міцно зв'язані з білком.
Флавінові дегідрогенази каталізують
реакцію дегідрування субстратів. Приєднання двох атомів водню перетворює
окиснену форму коферментів (ФМН чи ФАД) у відновлену (ФМНН2 чи ФАДН2).
Активною частиною флавінових коферментів є ізоалоксазинова циклічна система.
Обидва атоми водню (тобто 2 електрони і 2 протони) приєднуються до атомів азоту
ізоалоксазинового кільця.

Рис. 4. Перетворення окисненої форми ФМН
чи ФАД у відновлену ФМНН2 чи ФАДH2
Таким чином, флавінові коферменти
зв'язують обидва протони, тоді як нікотинамідні коферменти, як розглянуто вище,
зв'язують 1 протон, а другий залишається у середовищі.
ФМН служить простетичною групою
ферменту НАДН-дегідрогенази. Цей фермент, що знаходиться у внутрішній мембрані
мітохондрій, окиснює НАДН:
![]()
Флавінові ферменти з простетичною
групою ФАД каталізують відрив атомів водню від молекул субстратів, тобто є первинними дегідрогеназами:
![]()
До них відносяться сукцинатдегідрогеназа, яка каталізує окиснення
бурштинової кислоти у фумарову, ацил-КоА-дегідрогеназа
та ряд інших ферментів.
Убіхінон
Кофермент Q
(убіхінон, кофермент Q10) – жиророзчинний
кофермент, який знаходиться у мітохондріях еукаріотичних клітин. Ф. Крейн та К. Фолкерс у 1957-1958 встановили
хімічну формулу цієї сполуки. Кількість ізопренових одиниць буває різною
залежно від виду живих організмів. У клітинах організму людини знаходиться
убіхінон тільки з 10 ізопреновими одиницями. В організмі людини цей кофермент
синтезується з мевалонової кислоти та похідних фенілаланіну та тирозину (рис. 5).
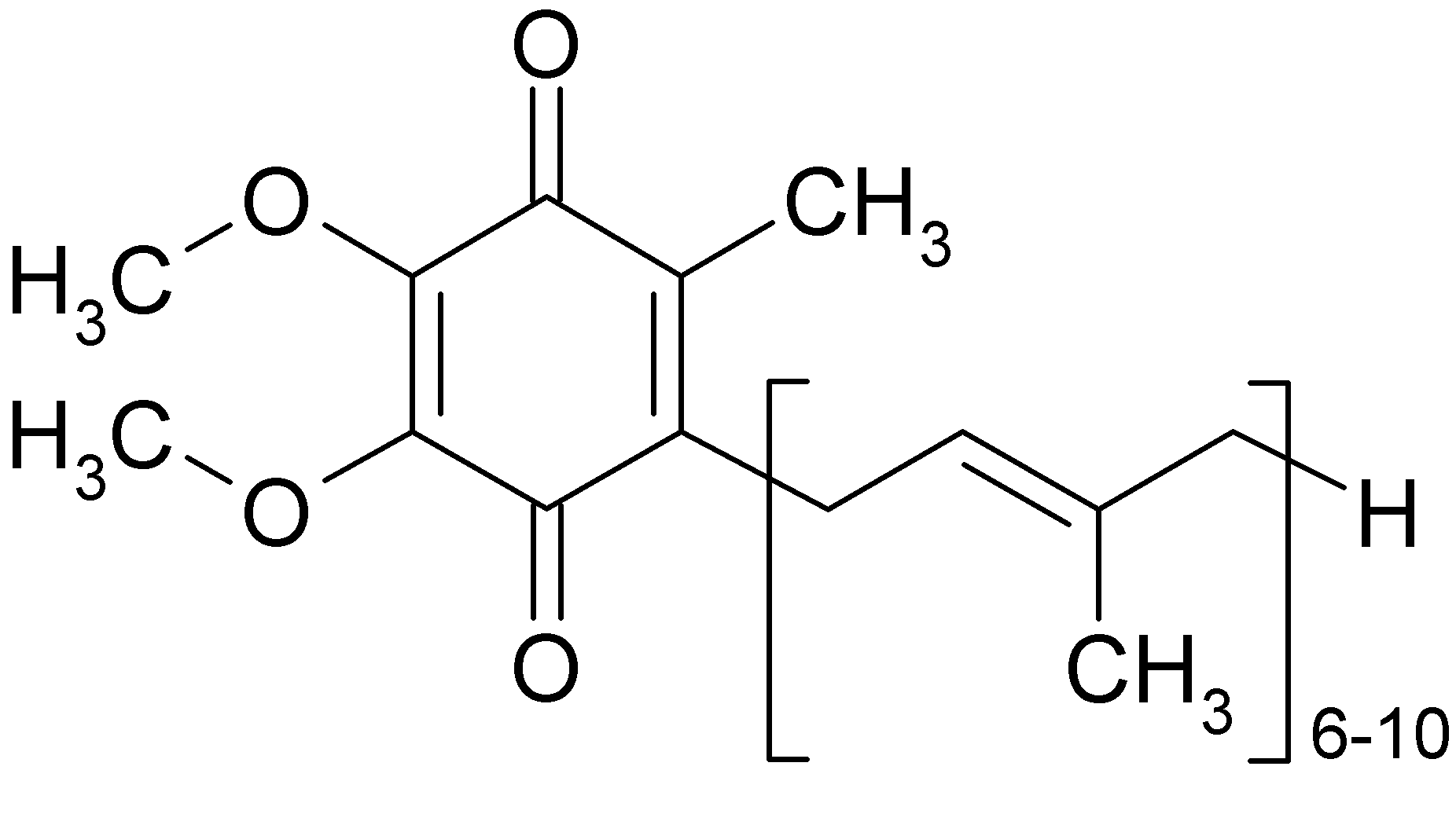
Рис.5 Хімічна формула убіхінону
Вміст убіхінону в тканинах залежить від інтенсивності енергетичного обміну.
Найбільша кількість коферменту Q у
серцевому м’язі.
|
Цікаво. Відомо, що у серцевому м’язі максимальний
вміст убіхінону спостерігається приблизно у 20 років, але потім з віком
поступово знижується: до 40 років – на 30%; після 60 років – на 50%. Саме
тому зміни функціональних можливостей серця в процесі старіння організму
пов’язують зі зниженням концентрації убіхінону у мітохондріях кардіоміоцитів.
Препарати коферменту Q10 використовують
з метою профілактики та у комплексному лікування серцево-судинних
захворювань. |
Убіхінон – це ліпофільна сполука, тому цей кофермент
розчинений у ліпідній частині внутрішньої мембрани мітохондрій, і, оскільки він
не зв’язаний з білком, може вільно рухатися в мембрані в різних напрямках.
Цей кофермент здатний приєднувати та віддавати
протони й електрони. Схема окисно-відновної реакції за участі убіхінону
наведена (рис. 6).



Рис.6. Реакція окиснення-відновлення убіхінону
У дихальному ланцюзі коензим Q є
колектором електронів від НАД- та ФАД(ФМН)-залежних дегідрогеназ.
Цитохроми
Цитохроми – складні
білки еммпротеїни, простетичною групою яких є гем. До складу гему цих
білків входить метал зі змінною валентністю (переважно залізо, може бути також
мідь). На сьогодні відомо приблизно 30 видів цитохромів.
Цитохроми вперше були описані Мак-Манном (Шотландія) у 1886 році й отримали
назву «гістогематини», але їх роль залишалася невідомою. У 1925 році
англійський біохімік Дейвід Кейлін почав активно вивчати ці білки. Насамперед було з’ясовано, що цитохроми
знаходяться у мембранах (мітохондріальних та ЕПР) всіх еукаріотичних клітин
(рис. 7).

Рис. 7. Структура гему у складі цитохрому b
Деякі цитохроми позначають цифровим індексом (наприклад, b5), який може бути
наданий тільки добре вивченим білкам.
У цитохромі с порфіринова площина, ковалентно зв’язана з білковою частиною через
два залишки цистеїну (рис. 8). В цитохромах b та а емм ковалентно не
зв’язаний з білком.

Рис. 8. Структура цитохрому с
У цитохромах аа3 замість
протопорфірину міститься порфірин А, який має певні структурні особливості. Усі
цитохроми мають різні фізико-хімічні властивості та величини стандартного
окисно-відновного потенціалу (редокс-потенціалу), що забезпечує напрямок руху
електронів від субстратів на кисень. Вони розташовані згідно з величинами їхніх
окисно-відновних потенціалів наступним чином: цитохром b, цитохром с1, цитохром с, цитохроми а, а3.
Цитохром аа3, або цитохромоксидаза, - це термінальный фермент, котрий переносить електрони
безпосередньо на кисень (аеробний фермент). Він складається з
6 субодиниць та, крім гемінового
заліза, містить іони міді.
Fe3+ ↔ Fe2+
У
перенесенні електронів спочатку беруть участь іони заліза
цитохромів а й а3, а далі - іон міді цитохрому а3.
Cu2+ ↔ Cu+
Унікальна особливість цитохрому аа3 полягає в тому, що цей
комплекс безпосередньо реагує з молекулярним киснем. Тому його також називають цитохромоксидазою. Молекула О2
зв'язується з атомом заліза цитохрому а3, після чого обидва атоми
кисню приймають по два електрони і, взаємодіючи з протонами, що надходять із середовища,
утворюють дві молекули води. Таким
чином, цитохромоксидаза каталізує чотирьохелектронне відновлення молекули О2
до води.
Залізо-сірчані
білки (FeS – білки)
Залізосірчані білки (FeS-білки)
- це білки з невеликою молекулярною масою (приблизно 10 кДа). Залізо, яке
вони містять не входить до складу гему, а зв’язане з атомами сірки (рис. 9).
Відомо, що вони беруть участь у транспорті електронів у дихальному ланцюзі, але
детально механізм окиснення-відновлення атомів заліза невідомий.

Рис. 9. Схема комплексу заліза і сірки в молекулі
залізо-сірчаного білка дихального ланцюга
Ці
білки локалізовані в ліпідному шарі мембран. Вони можуть зворотньо окиснюватися
та відновлюватися, тобто переносити електрони.
Подібні
окисно-відновні системи стабільні лише у складі білків. Вони можуть мати
від 2 до 6 іонів заліза, котрі
утворюють комплекси різного складу з неорганічним сульфідом і SH-групами залишків
цистеїну білкових молекул (рис. 10).

Рис. 10. Структура залізо-сірчаних
білків (FeS-білки)
Основні
компоненти дихального ланцюга
Мітохондрії – органели
клітини, функціонування яких для будь-якої обізнаної людини чітко асоціюється з
продукуванням енергії. Дійсно в матриксі мітохондрій локалізовані різноманітні ферменти,
що необхідні для окиснення субстратів. Крім того, внутрішня мембрана
мітохондрій містить систему білків-переносників електронів, які забезпечують
термінальний етап окиснення субстратів і створюють умови для синтезу АТФ (рис.
11).

Рис. 11. Структура мітохондрії
http://www.youtube.com/watch?v=-_8aYKcQZ_Q&feature=related

Рис. 12. Молекулярна організація
ланцюга перенесення електронів
http://www.youtube.com/watch?v=StXlo1W3Gvg&feature=related
Саме така послідовність компонентів не
є випадковою, а зумовлена величинами їх окисно-відновного потенціалу (Ео).
Ця константа кількісно характеризує здатність окисно-відновної пари, тобто
здатність окисненої і відновленої форм певної сполуки зворотно віддавати
електрон. Чим нижча (негативніша) величина ОВП пари, тим вища її можливість
віддавати електрони, тобто окиснюватися. І навпаки, пара з більш високим
(позитивним) значенням Ео буде приймати електрони і відновлюватись (табл. 1). Таким чином, електрони переходять від
однієї ОВ пари до іншої в напрямку більш позитивного Ео. Таке
перенесення електронів супроводжується зменшенням вільної енергії. Для ОВ
реакцій зміну стандартної вільної енергії визначають за рівнянням
![]()
де ![]() – зміна стандартної вільної енергії
реакції;
– зміна стандартної вільної енергії
реакції;
n – кількість перенесених
електронів (або атомів водню);
F – константа Фарадея (тепловий еквівалент
роботи, рівний 95 кДж);
∆Ео – різниця ОВ
потенціалів між двома парами.
Чим більша різниця ∆Ео
двох окисно-відновних пар, тим більше виділяється вільної енергії при
перенесенні електронів.
Табл. 1. Окисно-відновні
потенціали компонентів дихального ланцюга

Значення Ео в послідовності
від НАД+ до О2 поступово зростає. При повному переході
двох електронів від ОВ пари НАДН/НАД+ (Ео=‑0,32 В)
до ОВ пари Н2О/1/2О2 (Ео
=+0,82 В) зміна вільної енергії дорівнює 220 кДж (52,6 ккал). Кожний акт переходу
електронів між проміжними компонентами дихального ланцюга супроводжується
виділенням певної порції вільної енергії
(рис. 13).

Рис. 13. Вивільнення енергії в ланцюгу біологічного окиснення
Таким чином, завдяки наявності в ланцюгу
перенесення електронів від субстратів до О2 великої кількості
проміжних переносників енергія виділяється порціями і може бути використана для
синтезу декількох молекул АТФ. Синтез АТФ із АДФ і Фн у стандартних умовах
потребує 34,5 кДж/моль (8,3 ккал/моль), а в умовах живої клітини –
приблизно 50 кДж/моль (12 ккал/моль). Перепад енергії між НАДН/НАД+
і Н2О / 1/2О2 (220 кДж/моль)
достатній для синтезу не менше 4 молекул АТФ. Але експериментальні дослідження
показали, що синтезується максимум 3 молекули АТФ. Саме на 3 ділянках
дихального ланцюга перенесення електронів від одного компонента до наступного
супроводжується перепадом вільної енергії, достатнім для синтезу АТФ.
Усі учасники ланцюга перенесення електронів структурово об’єднані в
чотири окисно-відновні системи – мультиферменті комплекси I – IV.
Процес окиснення починається з перенесення
протонів і електронів з
субстрату, що окиснюється, на НАД+ або ФАД. Це залежить від природи субстрату. Кожен із комплексів
здатний каталізувати певну частину повної послідовності реакцій ланцюга (рис.
14).

Рис. 14. Структура дихального ланцюга
http://www.youtube.com/watch?v=3rO26W1xG9U&feature=related
Ці комплекси є
частиною внутрішньої мембрани мітохондрій.
http://www.youtube.com/watch?v=eizHVQfeMwo&feature=related
Комплекс I – НАДН-дегідрогеназа –
флавопротеїн, що містить ФМН. Цей фермент окиснює
НАДН і передає два атоми водню (2Н+ 2е-) на коензим
Q. Комплекс
також містить FeS-білки.
Комплекс II – сукцинатдегідрогеназа – флавопротеїн, що містить ФАД.
Цей фермент окиснює сукцинат і транспортує
два атоми водню (2Н+ 2е-) на
коензим Q. У складі комплекса присутні FeS-белки. У матриксі мітохондрій також містяться й інші ФАД-залежні дегідрогенази,
котрі окиснюють відповідні субстрати
(гліцерол-3-фосфат, ацил-КоА) та далі передають атоми водню на коензим
Q. Потоки
атомів водню об’єднуються на стадії утворення відновленого КоQН2 .
Коензим Q є останнім компонентом ланцюгу, котрий здатний транспортувати не тільки протони, але й електрони (2Н+ 2е-). Далі
протони (2Н+) переходять із
внутрішньої поверхні мембрани мітохондрії
на зовнішню, а електрони (2е-) крізь ланцюг цитохромів
переносяться на кисень.
Комплекс
III – убіхінондегідрогеназа – це ферментний комплекс, котрий
включає цитохром b, FeS-білок і цитохром с1. Цей комплекс транспортує електрони 2е-
від відновленого убіхінону КоQН2 на
цитохром с (невеликий за розмірами водорозчинний білок, що міститься на зовнішній стороні внутрішньої мембрани).
Комплекс IV – цитохром с-оксидаза – ферментний комплекс, що складається з цитохромів а й а3. Ці
ферменти
здійснюють останню стадію
біологічного
окиснення – відновлення електронами ( 2е- ) молекулярного кисню:
2е + 1/2О2 = О2-
Відновлений кисень О2- реагує з вільними протонами (2Н+
) матриксу. В результаті реакції утворюється ендогенна, або метаболічна вода (рис. 15):

Рис. 15. Утворення метаболічної води в результаті реакції
відновленого кисню О2, який реагує з вільними протонами 2Н+ матриксу
|
Напрямок перенесення протонів і електронів визначають
окисно-відновні потенціали. Для забезпечення
спонтанного перенесення компоненти
окисно-відновного ряду повинні розташовуватися згідно зі збільшенням величин
потенціалів. |
При проходженні по дихальному
ланцюгу пари електронів вивільняється енергія, більша частина котрої (60%) розсіюється у вигляді тепла,
а інша акумулюється у макроергічниз зв’язках АТФ,
а саме поглинається в реакції синтезу АТФ-окисному
фосфорилюванні (рис. 16) .

Рис. 16. Вивільнення енергії при проходженні по
дихальному ланцюгу пари електронів
Це фосфорилювання
називається окисним,
тому що енергія, що необхідна для
утворення макроергічного зв’язку, генерується у процесі окиснення, тобто руху
протонів і електронів по мітохондріальному ланцюгу транспорту електронів
Перша така ділянка – це
НАД ®ФМН, друга – цитохром b ®цитохром
с1, третя – цитохром аа3 ® кисень. Ці ділянки називають пунктами
фосфорилювання. Термін “пункт фосфорилювання” чи “ділянка фосфорилювання” не
треба розуміти як конкретну стадію, на якій безпосередньо відбувається
утворення АТФ. Ідеться про те, що потік електронів через ці три ділянки ланцюга
якимось чином поєднаний з утворенням АТФ (перепад ОВП тут достатній для
синтезу 1 молекули АТФ).
При окисненні субстратів ФАД-залежними
дегідрогеназами (наприклад, сукцинату сукцинатдегідрогеназою) потік електронів
від ФАДН2 до кисню не проходить через перший пункт фосфорилювання. У
цих випадках синтезується на 1 молекулу АТФ менше, тобто дві.
Механізм поєднання перенесення
електронів і
окиснювального фосфорилювання
Синтез АТФ із АДФ і Фн за рахунок енергії, що звільнюється при перенесенні
електронів по дихальному ланцюгу, називається окиснювальним фосфорилюванням
(ОФ). Існує декілька доказів поєднання цих процесів. По-перше, окиснення НАДН
чи ФАДН2 у мітохондріях дійсно супроводжується одночасним синтезом
АТФ. По-друге, при наявності інгібіторів
дихального ланцюга утворення АТФ зменшується. По-третє, відкриті хімічні речовини,
які роз'єднують процеси переносу електронів і синтезу АТФ, не порушуючи
перенесенні електронів по дихальному ланцюгу.
Тканинне дихання й ОФ поєднані не
тільки енергетично, а і просторово на внутрішній мембрані мітохондрій. У ній
локалізовані компоненти дихального ланцюга і ферменти, які каталізують реакцію
синтезу АТФ. Через зовнішню мембрану мітохондрій легко проникають майже всі
молекули й іони невеликих розмірів, тоді як внутрішня мембрана для більшості
іонів невеликого розміру є непроникною. У ній є спеціальні транспортні системи,
які переносять із цитоплазми в матрикс мітохондрій піруват та інше клітинне
паливо, а також АДФ і неорганічний фосфат, а з мітохондрій у цитоплазму – АТФ.
Матрикс містить піруватдегідрогеназну систему, ферменти циклу лимонної кислоти
(циклу Кребса), ферменти окиснення жирних кислот тощо. Саме НАД- та ФАД-залежні
дегідрогенази цих метаболічних шляхів переносять атоми водню на дихальний
ланцюг. Компоненти дихального ланцюга утворюють у внутрішній мембрані
високоупорядкований ансамбль. Точна локалізація всіх компонентів поки що не
вивчена, але встановлено, що деякі білки розміщені на одному боці внутрішньої
мембрани, а інші – на протилежному. Крім того, деякі білки дихального ланцюга
пронизують мембрану наскрізь. Фермент, який каталізує синтез АТФ (АТФ-синтаза
або Н+‑АТФаза), складається з двох компонентів: Fо
і F1 (рис.17).

Рис. 17. Схема окисного фосфорилювання за хеміосмотичною теорією
http://www.youtube.com/watch?v=YndC0gS3t6M&feature=related
Компонент F1 нагадує шапочку
гриба і повернений у бік матрикса мітохондрій. За допомогою ніжки він
прикріплений до компонента Fo, який фіксується у мембрані й пронизує
її наскрізь.
Існують три основні гіпотези про
механізм поєднання (спряження) дихання й ОФ: хімічна, хеміосмотична і
конформаційна. Згідно з хімічною гіпотезою, енергія, що виділяється при
перенесенні електронів по дихальному ланцюгу, використовується для утворення
високоенергетичного проміжного комплексу. Далі цей проміжний продукт
розщеплюється і передає енергію на утворення АТФ. Але всі спроби відкрити
проміжний високоенергетичний комплекс не дали результату.
Гіпотеза конформаційного поєднання
припускає, що виділення енергії при транспорті електронів зумовлює зміни
конформації білкових компонентів внутрішньої мембрани мітохондрій. Зміни
конформації передаються на АТФ-синтазу, що призводить до її активації.
Хеміосмотична теорія Мітчелла (теорія окисного
фосфорилювання)

П. Мітчелл
Пояснення механізмів спряження роботи дихального ланцюга та синтезу АТФ
було запропоноване англійським біохіміком Пітером Мітчеллом у 1961 році, яке
потім знайшло експериментальне підтвердження у роботах багатьох дослідників. За
винайдення механізмів окисного фосфорилювання П. Мітчелл у 1978 році отримав
Нобелівську премії в галузі хімії («За внесок у розуміння процесу перенесення
біологічної енергії, який зроблений завдяки створенню хеміосмотичної теорії»).
Основні постулати теорії Мітчелла:
1)внутрішня
мембрана мітохондрій непроникна для іонів та малих молекул (за винятком молекул
води);
2)дихальний
ланцюг працює як «помпа», що викачує протони з матриксу в міжмембранний простір
– рух 2 електронів від субстрату на кисень призводить до перенесення 8-10 Н+
(протони транспортуються через I, III
та IV комплекси) через мембрану;
3)робота
дихального ланцюга створює електрохімічний градієнт протонів (∆μН+),
оскільки вони вільно через внутрішню мітохондріальну мембрану повернутися в матрикс
не можуть і будуть накопичуватись у міжмембранному просторі; ∆μН+
- це проміжна форма зберігання енергії окиснення субстратів;
4)енергію
протонного градієнта використовує Н+-АТФ- синтаза (V
комплекс) для синтезу АТФ, коли через одну з її субодиниць протони повертаються
в матрикс;
5)існують
сполуки - роз’єднувачі окисного фосфорилювання, які порушують
електрохімічний градієнт протонів і знижують ефективність роботи Н+-АТФ-синтази.
На рис 18
наведена структура протонної АТФ-синтази (Н+-АТФ-синтази, Н+-АТФ-аза),
яка безпосередньо використовує енергію протонного градієнта, що був створений
дихальним ланцюгом, для синтезу АТФ.

Рис. 18. Структура Н+-АТФ-синтази
Протонна АТФ-синтаза – це олігомерний білок, який
вмонтований у внутрішню мембрану мітохондрії і за будовою нагадує гриб. Вона
містить дві субодиниці:
Fo – протонний канал (о - від «олігоміцин»); лише через
цей канал протони можуть повернутися в матрикс;
F1 – фермент, який
використовує енергію, що вивільняється при транспорті протонів через Fo для синтезу АТФ з АДФ та Фн.
П.
Мітчелл у своїй теорії теоретично віддав функцію спряження окиснення та
фосфорилювання саме Н+-АТФ-азі (рис. 19).

Рис. 19. Спряження роботи дихального ланцюга
із синтезом АТФ (за теорією
П.Мітчелла)
На сьогодні відомо, що при транспорті протонів через Fo-субодиницю
відбуваються конфірмаційні зміни в активному центрі F1-субодиниці,
які призводять до її активації і відповідно до синтезу АТФ та її вивільнення.
Молекули АТФ, що синтезуються, транспортуються у цитозоль за допомогою транслокази.
Для синтезу молекули АТФ, її вивільнення та
транспорту у цитозоль потрібна енергія 4 протонів (40% цієї енергії іде на
синтез АТФ, 60% виділяється у вигляді тепла). Кількість молекул неорганічного
фосфату, яке перейшло у зв’язану форму (тобто у АТФ) у перерахунку на один атом
кисню має назву коефіцієнт окисного фосфорилювання і позначається Р/O
(коефіцієнт фосфорилювання).
Коефіцієнт Р/O чисельно
дорівнює кількості молекул АТФ, що синтезувалися в результаті транспорту 2
ē на один атом кисню. Тому для субстратів, що окислюються під дією
НАД-залежних дегідрогеназ Р/O=3 (наприклад,
для пірувату, α-кетоглутарату, ізоцитрату, малату). Для субстратів, які
окислюються за допомогою ФАД-залежних дегідрогеназ, цей коефіцієнт дорівнює 2
(наприклад, для сукцинату, ацил-КоА, гліцерил-3-фосфату).
Згідно з цією
теорією, перенесення електронів по дихальному ланцюгу супроводжується
викачуванням протонів із матриксу через внутрішню мембрану у водне середовище
міжмембранного простору (рис. 20).

Рис. 20. Схема орієнтованого розташування оксидно-відновних петель,
які забезпечують перекачування протонів із матриксу в міжмембранний простір в
дихальному ланцюгу
Припускають, що асиметрично розміщені в
мембрані компоненти дихального ланцюга утворюють три петлі, які переносять
через мембрану протони, тобто служать протонними помпами. З кожною парою
електронів, що передаються від субстрату до кисню, ці три петлі транспортують
із матриксу мітохондрій шість протонів (за новими даними, не менше 9). Таким
чином, енергія, яка виділяється при перенесенні електронів, затрачається на
перекачування іонів Н+ проти градієнта концентрації. Внаслідок
викачування іонів Н+ із матриксу внутрішній бік внутрішньої мембрани
мітохондрій стає електронегативним, а зовнішній – електропозитивним, тобто виникає
градієнт концентрації іонів водню: їх менше в матриксі й більше – в
зовнішній водній фазі. Сумарний електрохімічний протонний потенціал
позначається ∆μН+. Він складається із 2-х компонентів: ∆μН=∆рН
і ∆V (рис. 21).

Рис. 21. Дві складові електрохімічного
протонного градієнта, що виникають
на внутрішній мітохондріальній мембрані
Внутрішня мембрана
мітохондрій непроникна для іонів Н+, а також іонів ОН-, К+,
Nа+, СI-, але мембранний
білок Fo АТФази утворює канал, по якому іони
Н+ повертаються в матрикс за градієнтом концентрації, вільна
енергія, яка при цьому виділяється, використовується F1-компонентом
АТФази для синтезу АТФ з АДФ і Фн.
http://www.youtube.com/watch?v=UL3M06aaMHM&feature=related
http://www.youtube.com/watch?v=StXlo1W3Gvg&feature=related
Таким чином, згідно з хеміосмотичною теорією, перенесення електронів по
дихальному ланцюгу створює градієнт концентраціїї іонів Н+, який
служить рушійною силою синтезу АТФ. Специфічний білок-переносник, аденіннуклеотид-транслоказа переносить
новосинтезований АТФ із матриксу в цитоплазму в обмін на АДФ. Ця сама
транспортна система переносить у матрикс мітохондрій фосфат.
Але найпереконливішим підтвердженням
гіпотези Мітчела став експеримент американського науковця Рекера: так званий дослід
із “химерою”. Назва “химера” запозичена зі старогрецької міфології про
чудовисько з пащею лева, тулубом кози і хвостом дракона.
За аналогією Рекер зібрав композицію з
трьох різних частинок. Спочатку він сконструював мембрану з фосфоліпідів сої, в
яку вмонтував бактеріальний білок родопсин, що має здатність у відповідь на
освітлення переносити протони проти градієнта концентрації (протонна помпа).
Третім компонентом мембранної композиції був фермент АТФ-синтетаза, одержаний
із міокарда бика. Таким чином, композиція мала всі компоненти, необхідні для
утворення АТФ. І справді, під впливом пучка світла "химера" ожила,
вона почала виробляти АТФ, незважаючи на всю неоднорідність складових частин.
Із цього часу хеміосмотична гіпотеза перетворилася на струнку теорію, за яку
Мітчелу в 1978 році було присуджено Нобелівську премію.
Шляхи використання АТФ в організмі:
Енергія гідролізу АТФ використовується
клітинами для виконання всіх відомих ендергонічних процесів (рис. 22):
1)
реакцій синтезу вуглеводів, ліпідів, білків, нуклеїнових кислот;
2) механічної роботи, наприклад скорочення
м'язів, руху хромосом при мітозі;
3) активного перенесення речовин через
мембрани проти градієнта концентрації;
4) забезпечення точної передачі генетичної
інформації;
5) електричної роботи – проведення нервового
імпульсу.

Рис.
22. Шляхи використання енергії
АТФ
Отже, АТФ виступає
як переносник хімічної енергії, який зв'язує клітинні процеси, що супроводжуються
виділенням енергії, з тими головними видами клітинної активності, в яких
енергія споживається.
Регуляція тканинного дихання та
окисного фосфорилювання
Швидкість
окиснення субстратів клітинного палива і транспорту електронів до кисню по дихальному
ланцюгу залежить від наявності АДФ і Фн та виражається рівнянням:
НАДН + Н+ + 1/2 О2 + 3 АДФ + 3 Фн ® НАД+
+ 3 АТФ +4 Н2О
Концентрація АДФ у клітині значно
менша, ніж концентрація неорганічного фосфату, тому рівень АДФ є основним у регуляторному
механізмі. Якщо в клітині інтенсивно здійснюються процеси з використанням
енергії АТФ, то це призводить до зниження концентрації АТФ і зростання
концентрації АДФ, а наявність АДФ автоматично підвищує швидкість перенесення
електронів та поєднаного з ним окиснювального фосфорилювання. Після вичерпання
запасів АДФ швидкість споживання мітохондріями кисню і фосфорилювання АДФ
неодмінно зменшуються, повертаючись до рівня, що відповідає стану спокою.
Залежність дихання мітохондрій від концентрації АДФ називають дихальним
контролем. Таким чином, тісне поєднання дихання з фосфорилюванням у
мітохондріях створює умови, коли швидкість окиснення клітинного палива регулюється
енергетичними потребами клітини. Величина відношення АТФ/АДФ•Фн служить
показником енергетичного стану клітин.
Запропонований ще один показник –
енергетичний заряд, який відображає вміст високоенергетичних аденілових
нуклеотидів у загальному їх пулі:
АТФ +
1/2 АДФ
енергетичний заряд =
—————————
АТФ +
АДФ + АМФ
Більшість клітин підтримують значення
енергетичного заряду у вузькому інтервалі близько 0,8-0,9. АТФ, АДФ і АМФ
відіграють роль позитивних або негативних алостеричних ефекторів регуляторних
ферментів метаболізму вуглеводів, жирів, амінокислот. При високій концентрації
АТФ і відповідно низьких концентраціях АДФ і АМФ процеси катаболізму й
окиснювального фосфорилювання мінімальні. З іншого боку, підвищення рівня АДФ і
АМФ є сигналом, який збільшує швидкість катаболізму і зменшує швидкість
анаболізму.
При збільшенні в клітині споживання АТФ поідвищується
швидкість надходження АДФ в мітохондрії,
що означає зростання швидкості синтезу АТФ. Це призводить до збільшення
швидкості транспорту протонів: зростає швидкість дихання. Така кореляція, а саме пришвидшення окисного фосфорилювання та дихання
при підвищенні концентрації АДФ називається дихальним контролем.
Дихальний контроль
Дихальний контроль – це співвідношення АТФ/АДФ: При його
зменшенні підвищується інтенсивність дихання. Процес синтезу
АТФ залежить від енергетичного статусу клітини, який, у свою чергу знаходить
своє вираження у кількісному співвідношенні АТФ/АДФ. При зростанні потреб клітини в
енергії підвищується концентрація АДФ і значення цього співвідношеня
знижується. Це супроводжується активацією тканинного дихання і, відповідно,
окисного фосфорилювання.
Процес залежності дихання
клітин від вмісту аденілових нуклеотидів був вивчений in vitro.
Результати дослідження показали, що за наявності в середовищі всіх компонентів,
за винятком АДФ, поглинання кисню клітиною не відбувається. Внесення в
інкубаційну суміш АДФ стимулює дихання, активність якого знижується при
зростанні концентрації АТФ. Швидкість дихання знижується до нуля, коли вся АДФ
перетворюється в АТФ. Тому залежність роботи дихального ланцюга від співвідношення АТФ/АДФ отримала
назву дихальний контроль. Механізм дихального контролю відрізняється
високою точністю і швидко реагує навіть на невеликі зміни концентрацій АТФ та
АДФ. Незначне підвищення концентрації АДФ призводить до активації дихання та
фосфорилювання. Зниження синтезу
АТФ призводить до розвитку гіпоенергетичного стану в організмі.
Синтез АТФ
знижується при:
1)гіпоксії;
2)голодуванні,
дефіциті деяких вітамінів (В1, В2, РР) та мінералів (Fe, Cu);
3)наявності
інгібіторів дихального ланцюга та інгібіторів окисного фосфорилювання;
4)наявності
роз’єднувачів окиснення та фосфорилювання.
Інгібітори та роз’єднувачі окисного фосфорилювання
Робота
протонної АТФ-синтази може бути порушена за наявності в клітині сполук – роз’єднувачів
окисного фосфорилювання. Так, наприклад, олігоміцин пригнічує роботу
Fo субодиниці і, таким чином, робить неможливим
використання енергії протонного градієнта для синтезу АТФ.
Роз»єднувачі – це сполуки ліпофільні, легко дифундують через внутрішню
мембрану мітохондрій, можуть зв’язувати протони та переносити їх у матрикс за
градієнтом концентрацій Н+ (тому вони належать до сполук
протонофорів).
У разі наявності в клітині роз’єднувача окиснення субстратів відбувається
дуже активно, дихальний ланцюг мітохондрій працює і намагається створити
електрохімічний градієнт протонів, але цього не відбувається. Пояснення в тому,
що молекули роз’єднувача захоплюють протони з міжмембранного простору і
транспортують їх у матрикс. За таких умов окиснення субстратів не приводить до
створення протонного градієнта і, відповідно, до синтезу АТФ. Енергія окиснення
перетворюється в теплову енергію, тобто коефіцієнт окисного фосфорилювання
знижується і спостерігається виділення тепла (пірогенна дія).
Прикладом роз’єднувача служить 2,4-динітрофенол. Механізм токсичної дії
2,4-динітрофенолу полягає у блокуванні окиснювального фосфорилювання, що
викликає утворення енергії шляхом гліколізу з використанням значної кількості
кисню. Оскільки вказана
енергія не накопичується в макроенергічних сполуках фосфору, вона виділяється у
вигляді тепла, викликаючи гіпертермію.

Рис. 23. Структурна формула роз’єднувача
2,4-динітрофенолу
Крім 2,4-динітрофенолу до синтетичних роз’єднувачів
потрібно віднести деякі похідні вітаміну К, анестетики, антибіотики валіноміцин
та граміцидин, ацетилсаліцилову кислоту, гербіциди. До природних роз’єднувачів
належать продукти перикисного окиснення ліпідів, довголанцюгові жирні кислоти,
великі дози териоїдних гормонів.
Саме тому в людей, які хворіють на
гіпертиреоз спостерігається підвищення температури тіла, активне окиснення
субстратів і тому, незважаючи на підвищений апетит, вони втрачають вагу. Відомо
також, що в разі купання у холодній воді в організмі людей-моржів
спостерігається підсилена секреція тиреоїдних гормонів, що, у свою чергу,
дозволяє підсилити термопродукцію і покращити терморегуляцію.
Прикладом природного роз’єднання також може бути робота білка термогеніну,
який знаходиться у внутрішній мембрані мітохондрій бурої жирової тканини.
Цей білок є протонним каналом, через який транспортуються протони у матрикс
і тому робота дихального ланцюга не може створити необхідний електрохімічний
градієнт протонів для синтезу АТФ (рис. 24). Тому активне окиснення субстратів
бурою жировою тканиною призводить до генерації теплової енергії.

Рис. 24. Механізм дії природного роз”єднувача
термогеніну
|
Цікаво. Бура жирова тканина добре розвинена у новонароджених дітей. У
дорослих вона залишається в незначній кількості на шиї, між лопатками,
навколо нирок, щитоподібної залози та аорти. Бура жирова тканина містить
велику кількість мітохондрій, цитохроми яких надають тканині коричневого кольору. Мітохондрії цієї жирової тканини
мають окислювальну здатність, що у 20 разів більша, ніж мітохондрій білої
жирової тканини. Для дітей ця тканина має велике значення, оскільки невеликий за
розмірами організм дитини здатний дуже швидко втрачати тепло. Кількість
активного бурого жиру у дорослих різна і залежить від ваги. Як правило, у
гладких людей кількість бурої жирової тканини менша. Організм дорослих, які
мають буру жирову тканину, відрізняється підвищеною швидкістю метаболізму,
оскільки, як відомо, |
Велика кількість речовин, у тому числі й
лікарські засоби, можуть змінювати енергетику клітин, впливаючи на утворення
енергії в ході перетворення поживних речовин і на рівень окислювального
фосфорилювання (утворення АТФ). Їх можна розділити на активатори й інгібітори
енергетичного обміну.
До активаторів відносяться: глюкоза,
фруктоза, амінокислоти, кислоти циклу Кребса (лимонна, яблучна, янтарна), різні
поживні суміші, наприклад препарати гідролізатів білків (гідролізат казеїну). Вони поліпшують енергетичний обмін у
тканинах організму, залучаючись до окиснювально-відновних реакцій мітохондрій,
і тому знайшли застосування в медичній практиці.
Інгібітори знайшли застосування в наукових дослідженнях під час вивчення
окремих етапів обміну речовин та енергії.За механізмом дії речовини, які впливають
на енергетичний обмін у клітинах, можна поділити на чотири групи.
Інгібітори дегідрогеназ - гальмують
процес дегідрування окремих субстратів, знижуючи надходження атомів водню
(протонів і електронів) у дихальний ланцюг. До них належать, наприклад,
протитуберкульозні препарати фтивазид, ізоніазид (ГІНК) та інші, які є
похідними ізонікотинової кислоти:

Ізонікотинова
Ізоніазид (гідразид)
Препарати
ізоніазид, фтивазид, салюзид та інші, як похідні ізонікотинової кислоти, є структурно
подібними до аміду нікотинової кислоти й тому виступають конкурентними
інгібіторами нікотинамідних дегідрогеназ (НАД-залежних), які містять у складі
своїх коферментів амід нікотинової кислоти. Тому має місце їх конкурентне
заміщення, яке призводить до пригнічення дії НАД-залежних дегідрогеназ, і
механізм клітинного дихання в мікроорганізмів пригнічується, що веде до їх
загибелі.
Малонова кислота (Н00С-СН2-С00Н)
- нормальний проміжний продукт обміну - є конкурентним інгібітором ФАД-залежної
дегідрогенази (сукцинатдегідрогенази), яка відщеплює атоми водню від янтарної
кислоти (Н00С-СН2-СН2-С00Н) - одного із субстратів циклу
Кребса, у зв'язку з чим швидкість цього циклу знижується. Суміш інгібіторів
НАД- і ФАД-залежних дегідрогеназ може значно пригнічувати тканинне дихання, не
впливаючи на утворення протонного потенціалу, оскільки надходження іонів Н+
з матриксу назовні забезпечує убіхінон.
Інгібітори
тканинного дихання на етапах фосфорилювання блокують одну з трьох ланок
утворення АТФ, перепиняючи потік електронів на певних ділянках дихального
ланцюга. Перша група препаратів (снотворні препарати барбітурового ряду -
амітал та ін., прогестерон - жіночий статевий гормон) припиняє надходження
водню на дихальний ланцюг від субстратів, які окиснюються внаслідок дії
піридинзалежних дегідрогеназ, але не заважають використанню субстратів, які
окиснюються через ФАД (наприклад, янтарної кислоти).
На рівні другої ланки дихальний
ланцюг блокується протигрибковим антибіотиком антиміцином А (блокується перенос
електронів між цитохромами b і с,). Дихання можливе тільки в разі надходження
електронів і протонів на ділянку ланцюга після блоку. Наприклад, аскорбінова
кислота (вітамін C) може окиснюватися в цитохромом с. Тому в її присутності
дихання в мітохондріях продовжується, незважаючи на те, що дихальний ланцюг
гальмується антиміцином А.
Третя група інгібіторів дихання
(ціаніди (NaCN, KCN), азиди (NaN3), оксид вуглецю (II) та ін.) блокують
цитохромоксидазу й унеможливлюють сам процес дихання. Ці речовини викликають кисневе
голодування для дихального ланцюга мітохондрій, хоча кисень наявний у великій
кількості (блокується процес перекидання електронів на кисень). Тому
вимикається утворення протонного потенціалу й поєднане з ним фосфорилювання -
припиняється життєдіяльність клітин. Перелічені інгібітори цитохромоксидази є
найсильнішими отрутами, отруєння якими викликає швидку загибель організму (рис.
25).

Рис. 25. Інгібітори тканинного дихання
Іонофори (переносники іонів). До них належать речовини, здатні зв'язувати
певні іони (К+, та ін.) і переносити їх через мембрани, порушуючи їхній
ізолюючий бар'єр. Від роз'єднуючих агентів вони відрізняються тим, що
переносять через мембрану не іони Н+, а будь-які інші катіони. Наприклад,
токсичний антибіотик валіноміцин утворює жиророзчинний комплекс з іонами К+,
який легко проходить через внутрішню мембрану мітохондрій, тоді як за
відсутності валіноміцину іони К+ проникають крізь неї з великими труднощами.
Антибіотики-іонофори вирівнюють
іонні градієнти на будь-якій, а не лише на мітохондріальній мембрані, тому,
припиняючи вироблення енергії й вирівнюючи іонні градієнти між внутрішнім і
позаклітинним середовищем, вони призводять до швидкої загибелі мікроорганізмів.
Іонофор граміцидин є антибіотиком з бактеріостатичною й бактерицидною дією. Він
полегшує проникнення крізь мембрану К+ і Na+, причому діє на клітини як
мікроорганізмів, так і хворого, тому його необхідно застосовувати тільки у вигляді
мазей і паст для лікування гнійних ран, остеомієлітів та у вигляді промивань і
полоскань при запальних захворюваннях вуха, горла тощо.
Інші види біологічного
окиснення
Розглянутий вище
різновид біологічного окиснення, при якому кисень використовується на утворення
води та вироблення енергії в дихальному ланцюзі, називається оксидазним, або
енергозабезпечувальним. На нього припадає 80-90 % спожитого кисню. Крім цього,
існує ще кілька видів реакцій, які відбуваються із споживанням кисню. Вклад їх у загальне споживання кисню
організмом порівняно невеликий (приблизно 10 %), але значення важливе. У
більшості реакцій атоми кисню безпосередньо включаються в молекулу субстрату з
утворенням гідроксильної групи, рідше – карбоксильної й інших кисневмісних груп.
Ці реакції зустрічаються на різних шляхах синтезу і розпаду, особливо в
метаболізмі ароматичних і стероїдних сполук, простагландинів і лейкотрієнів.
Крім того, реакції окиснення мають значення для метаболізму лікарських
препаратів, знешкодження сторонніх речовин. Найчисленнішими є реакції
мікросомального окиснення. Назва походить від мікросом – фрагментів мембран
ендоплазматичного ретикулуму у формі везикул, які утворюються при гомогенізації
тканин.
Мікросомальне окиснення
У мембранах ендоплазматичного ретикулуму печінки, в мітохондріях і
мікросомах кори надниркових, статевих залоз та інших тканин локалізовані
ферментні системи, які каталізують монооксигеназні реакції, коли один атом
молекули кисню включається в субстрат, а другий – у молекулу води. Оскільки найчастіше субстрат у
монооксигеназних реакціях гідроксилюється, цю групу ферментів називають також
гідроксилазами. Загальне рівняння реакцій мікросомального гідроксилювання таке:
RН + О2 + НАДФ Н + Н+
® RОН +
Н2О + НАДФ+
Донором воднів для утворення Н2О
замість НАДФН може бути НАДН, ФМН•Н2, ФАД•Н2.
Головний компонент монооксигеназ –
цитохром Р-450 – названий так тому, що комплекс його відновленої форми з
монооксидом вуглецю (II) має максимум поглинання світла при 450 нм. Цитохром
Р-450 містить протогем і подібний до цитохромів групи b. Буква “Р” в цитохромі
Р-450 походить від американського міста Philadelphia, де він вперше був
відкритий. Існує велика кількість ізоформ цитохрому Р-450. Функціонують
цитохроми Р-450 спільно з флавопротеїнами і залізо-сірчаними білками. Зокрема,
гідроксилазна система кори надниркових залоз включає цитохром Р-450,
залізо-сірчаний білок адренодоксин і флавопротеїн, коферментом якого служить
ФАД. Вона каталізує реакції гідроксилювання у процесі синтезу стероїдних
гормонів кори надниркових залоз. Послідовність компонентів ланцюга переносу
електронів показана на рис. 26.

Рис. 26. Схема монооксигеназної системи кори надниркових залоз
Флавопротеїн окиснює НАДФН і далі передає
електрони в ланцюг, а протони в середовище. В активному центрі цитохрому Р-450
зв'язуються О2 і субстрат, молекулярний кисень активується шляхом
перенесення на нього електронів і відбувається гідроксилювання субстрату
активним киснем. Другий атом кисню зв'язує два іони Н+ із
середовища, утворюючи молекулу води. Монооксигеназна система мембран
ендоплазматичного ретикулуму печінки включає флавопротеїн НАДФН-цитохром
Р-450-редуктазу і цитохром Р-450.
Вона каталізує реакції гідрокислювання у процесі синтезу жовчних кислот із
холестерину, інактивації стероїдних гормонів, метаболічні перетворення
різноманітних сторонніх речовин і лікарських препаратів. У результаті
гідроксилювання неполярних гідрофобних речовин підвищується їх гідрофільність,
що сприяє інактивації біологічно активних речовин чи знешкодженню токсичних
речовин і виведенню із організму. Ізоформи цитохрому Р-450 розрізняють за субстратною специфічністю, але
всі субстрати повинні бути аполярними. Субстрати мікросомальних гідроксилаз
індукують синтез молекул цитохрому Р-450 в печінці. Висока індукуюча здатність
властива фенобарбіталу (снодійному препарату). Явище індукції пояснює механізм
звикання до снодійних засобів (барбітуратів) і до інших лікарських речовин, які
окиснюються монооксигеназною системою ендоплазматичного ретикулуму гепатоцитів.
Існують монооксигенази, які не містять
цитохрому Р-450. Серед них ферменти печінки, що каталізують реакції
гідроксилювання фенілаланіну, тирозину, триптофану. До їх складу входять: іон
заліза й органічний фактор біоптерин. Мідьвмісна монооксигеназа каталізує
гідроксилювання дофаміну з утворенням норадреналіну.
Пероксидазне окиснення
Відомі реакції безпосереднього
окиснення субстратів киснем не шляхом включення атомів кисню в молекули субстратів,
а шляхом двохелектронного відновлення кисню до пероксиду водню. Це так звані
пероксидазні реакції. Вони перебігають за схемами:
RH2 + О2 ®R + Н2О2 або
RH +
О2 + Н2О ®RОН + Н2О2
Таким чином, продуктами цих реакцій є
окиснений субстрат та Н2О2. До ферментів, які каталізують
такі реакції, відносяться оксидази D- і L‑амінокислот, аміноксидази,
ксантиноксидази тощо. Пероксид водню далі розкладається каталазою або
використовується у реакціях, які каталізуються пероксидазами. Оксидази,
каталаза і пероксидази локалізовані, головним чином, у пероксисомах. Вони
містять флавін або метали.
Таким чином, крім використання кисню в
тканинному диханні у мітохондріях, в організмі широко розповсюджені реакції
прямої взаємодії субстратів з молекулярним киснем. Принципова відмінність цих
реакцій біологічного окиснення від тканинного дихання в тому, що вони не
супроводжуються утворенням високоенергетичних сполук, а мають пластичне і
захисне значення.
Вільнорадикальне окиснення
Реакції цього шляху відбуваються під дією активних форм кисню (АФК), які
містять неспарений електрон і тому є високореакційноздатними –
можуть окиснювати різноманітні сполуки в організмі і тим самим пошкоджують їх
структуру. АФК є агресивними молекулами по відношенню до білків, нуклеїнових
кислот і, що саме головне, до ліпідів біомембран. Вони запускають ланцюговий
процес перекисного окиснення ліпідів (ПОЛ), що призводить до руйнації
мембранних структур (рис 27).

Рис. 27. Схема утворення активних форм кисню в
НАДФ.Н-залежного
електронно-транспортного ланцюга фагоцитуючих клітин крові
АФК утворюються як бічні продукти роботи дихального ланцюга при неповному
відновленні кисню. Повне відновлення кисню супроводжується акцепцією 4
електронів і утворенням ендогенної води. В разі, коли кількість електронів, які
транспортуються на кисень, менша, утворюються АФК:
![]()
Крім того, ці форми кисню, насамперед супероксидний аніон радикал, можуть
утворюватися при функціонуванні деяких ферментів (рис. 28).

Рис. 28. Утворення, знешкодження та використання
АФК:
СОД – супероксиддисмутаза; МПО –
мієлопероксидаза.
Відомо, що 2-5% кисню, який поглинає організм, перетворюється саме в
ці активні молекули, найбільш реакційно здатною з яких є гідроксильний радикал
– він може окиснити будь-яку молекулу в організмі.
Утворення NO радикала (NO∙) та
подальша його взаємодія з супероксидним аніон радикалом призводять до утворення
пероксинітриту (ONOO-), який у
незначній кількості може виконувати функцію вазодилятатора, а при високій
концентрації викликає ушкодження клітин.
|
|

Утворення АФК відбувається постійно в клітинах організму і є звичайним
метаболічним процесом. Постійним джерелом АФК є макрофаги, які використовують
супероксидний радикал для боротьби із чужорідними організмами (вірусами,
бактеріями, злоякісними клітинами).
Вільнорадикальне окиснення є необхідним для утворення біорегуляторів
(простагландинів, лейкотрієнів, тромбоксанів) з арахідонової кислоти.
Лише 2% АФК використовується організмом у процесах,
корисних для організму. Решта (98%) АФК має руйнівну дію і може пошкоджувати
різні клітинні структури.
Негативний вплив АФК пов'язаний з їх здатністю
стимулювати утворення органічних пероксидів (ROOH). Цей
процес отримав назву перекисне окиснення. АФК найбільш активно атакують ненасичені
жирні кислоти, що знаходяться у складі фосфоліпідів біомембран. Як було
зазначено вище, найбільшу окиснювальну здатність має гідроксильний радикал, і
саме тому він ініціює ланцюг
вільнорадикального окиснення. Гідроксильний радикал забирає водень у СН2-групи
поліненасиченої жирної кислоти у складі
фосфоліпіду і, таким чином, утворюється ліпідний радикал (L∙).
Розвиток ланцюга відбувається шляхом приєднання кисню:
L • + О2
→ LOO •
LOO• + LH → LOOM + LR•.
В результаті утворюються ліпідний пероксирадикал LOO• та пероксид ліпіду LOOH. Радикали, що утворюються,
мають високу реакційну здатність – кожний радикал здатний генерувати утворення
декілька нових радикалів. Таким чином, виникає ланцюгова реакція перекисного
окиснення ліпідів.
Ланцюг вільнорадикального окиснення може бути обірваний як за рахунок
взаємодії радикалів один з одним, так і в реакціях із компонентами
антиоксидантної системи клітини (наприклад, вітаміном Е):
LOO• + L• →
LOOH + LH;
L• + вітамін E → LH +
вітамін E•;
вітамін E• + L• →
LH + вітамін Еокисл.
Накопичення АФК та активація перекисного
окиснення призводять до низки негативних наслідків у клітині:
1)порушенням рідинно-кристалічної структури ліпопротеїнів мемебран;
2)зниження міцності та порушення структури мембран клітини, в тому числі
мітохондріальної мембрани;
3)структурно-функціональні порушення ферментативних систем дихання та
оксидоредуктаз;
4)порушення ферментативних систем синтезу АТФ;
5)ушкодження транспортних мембранних систем;
6)гальмування процесів синтезу білків, нуклеїнових кислот, тощо;
7)руйнація лізосом та вивільнення лізосомальних гідролітичних ферментів;
8)накопичення продуктів неповного окиснення субстратів з можливим
подальшим розвитком ацидозу;
9)інактивація деяких коферментів (глутатіону, ліпоєвої кислоти).
http://www.youtube.com/watch?v=i3WZ2pyI-qM&playnext=1&list=PLD9620432F9A4D353
Антиоксидантна система (АОС) – це потужний механізм, що
запобігає розвитку вільнорадикальних та перекисних реакцій в організмі. Ця система
клітин організму діє завдяки наявності сполук - антиоксидантів, у складі яких
міститься рухливий атом водню, що не дуже міцно з’єднаний з вуглецем (С-Н) або
сіркою (S-Н). У результаті реакцій молекул
антиоксидантів та вільних радикалів утворюються радикали антиоксидантів, які не
є потужними окисниками й не можуть продовжувати перебіг вільнорадикальних
реакцій окиснення, тобто вони обривають ці ланцюги.
Антиоксиданти можуть знешкоджувати вільні радикали
ще до моменту реалізації їх руйнівної дії. Таким чином основним завданням
антиоксидантної системи є зменшення кількості вільних радикалів до мінімально
можливого рівня.
Усі
компоненти антиоксидантної системи організму умовно поділяють на декілька груп:
1)
ферменти АОС;
2)
макромолекулярні неферментні сполуки;
3)
низькомолекулярні неферментні сполуки
(жиророзчинні та водорозчинні антиоксиданти).