ОСОБЕННОСТИ ГЕНЕТИКИ ЧЕЛОВЕКА. ПРОЯВЛЕНИЯ ОСНОВНЫХ ЗАКОНОМЕРНОСТЕЙ НАСЛЕДОВАНИЯ НА ПРИМЕРЕ МЕНДЕЛИРУЮЩИХ ПРИЗНАКОВ ЧЕЛОВЕКА (МОНО -, ДИ - И ПОЛИГИБРИДНОЕ СКРЕЩИВАНИЕ). МНОЖЕСТВЕННЫЙ АЛЛЕЛИЗМ. ГЕНЕТИКА ГРУПП КРОВИ. ВЗАИМОДЕЙСТВИЕ АЛЛЕЛЬНЫХ И НЕАЛЛЕЛЬНЫХ ГЕНОВ. ЯВЛЕНИЕ ПЛЕЙОТРОПИИ.
Генетика человека изучает явления наследственности и изменчивости у человека на всех уровнях его организации и существования: молекулярном, клеточном, организменном и популяционном. Современная генетика человека базируется на законах классической генегики, которые имеют универсальное значение. Так же, как в классической генетике, появление и становление которой связано с изучением наследования мутационных изменений в популяциях гороха, дрозофилы, мыши и других экспериментальных объектов исследования, основные достижения в генетике человека обусловлены анализом природы и характера наследования мутационных изменений у человека.

ДНК
В последние годы выявлено, что спонтанная наследственная изменчивость весьма высока - в течение жизни человека приблизительно у 70% людей реализуются те или иные наследственные болезни. Таким образом, у большинства людей в течение их жизни проявляется хотя бы одно серьезное генетически обусловленное отклонение от нормы, снижающее продолжительность жизни человека по сравнению с нормой либо мешающее его нормальной жизнедеятельности и работоспособности. Изучение молекулярной природы таких генетических изменений, анализ закономерностей их наследования, оценка их распространенности в различных популяциях человека, изучение роли мутагенных факторов окружающей среды в возможном изменении спонтанного уровня мутагенеза у человека относятся к наиболее важным направлениям исследований в области генетики человека. Опираясь на эти фундаментальные знания, медицинская генетика разрабатывает методы диагностики, лечения и профилактики наследственной патологии, связанной с широким спектром менделевских, хромосомных и мультифакториальных наследственных болезней.
Современный этап развития генетики человека характеризуется стремительным прогрессом наших знаний о молекулярном строении генетического материала и о механизмах мутагенеза. Наглядным примером прогресса в области генетики человека являются успехи реализации международной программы "Геном человека". Интенсивное изучение наследственных болезней в клиниках многих стран увеличило к 1998 г их число почти до 9000 (в 1966 г было изучено только около 1500 наследственных болезней). Для более чем 3900 из этих недугов изучена локализация мутантных генов в хромосомах и проведен молекулярный анализ продуктов их деятельности. Эти достижения поставили на новую основу разработку методов диагностики наследственных болезней, их профилактики и генотерапии.

Близнецовый метод
В связи с возрастающим загрязнением окружающей среды, особенно связанным с радиационным загрязнением целых регионов в результате Чернобыльской аварии, ядерных взрывов на полигонах и деятельностью предприятий ядерного топливного цикла, важное значение приобретает разработка методов оценки генетических последствий такого загрязнения для грядущих поколений (Шевченко В.А. и др., 2004).
Успехи генетики человека, ее история, тесно связаны с развитием всех разделов генетики. Задолго до открытия Г. Менделя различными авторами были описаны патологические наследственные признаки у человека и основные типы наследования. Первые сведения о передаче наследственной патологии у человека содержатся в Талмуде (4 в. до н.э.), в котором указано на опасность обрезания крайней плоти у новорожденных мальчиков, старшие братья которых или дяди по материнской линии страдают кровотечением.
К XVIII в. относятся первые описания доминантного (полидактилии, т.е. шестипалости) и рецессивного (альбинизма у негров) признаков, сделанных французским ученым П. Мопертюи. В начале XIX в. несколькими авторами одновременно было описано наследование гемофилии, в результате изучения родословных семей, в которых встречались лица, страдающие этой болезнью.
В 1814 г. вышла книга лондонского врача Д. Адамса "Трактат о предполагаемых наследственных свойствах болезней, основанный на клиническом наблюдении." Позже она была переиздана под названием "Философский трактат о наследственных свойствах человеческой расы". Этот труд стал первым справочником для генетического консультирования. В нем Адаме сформулировал многие важные принципы медицинской генетики: "Браки среди родственников повышают частоту семейных (т.е. рецессивных) болезней", "Наследственные (доминантные) болезни не всегда проявляются сразу после рождения, но могут развиваться в любом возрасте", "Не все врожденные болезни являются наследственными, часть из них связана с внутриутробным поражением плода (например, за счет сифилиса)".
В середине XIX в. в России над проблемами наследственных болезней и наследственной природы человека работал В.М. Флоринский. В 1866 г. вышла его книга "Усовершенствование и вырождение человеческого рода". Наряду с противоречивыми или неверными положениями, в ней был поднят и правильно освещен ряд вопросов медицинской генетики. Среди них: значение среды для формирования наследственных признаков, вред близкородственных браков, наследственный характер многих патологий (глухонемоты, альбинизма, заячьей губы, пороков развития нервной трубки). Однако этот труд В.М. Флоринского не был оценен в полной мере его современниками в силу неподготовленности к восприятию этих идей.
В последней четверти XIX в. весомый вклад в развитие генетики человека внес английский биолог Ф. Гальтон, названный К.А. Тимирязевым "одним из оригинальнейших ученых, исследователей и мыслителей." Гальтон впервые поставил вопрос о наследственности человека как предмете для изучения наследственных признаков. Анализируя наследственность ряда семей, Гальтон пришел к выводу, что психические особенности человека обусловлены не только условиями среды, но и наследственными факторами. Кроме того, он предложил и применил близнецовый метод для изучения соотносительной роли среды и наследственности в развитии признаков. Им же разработан ряд статистических методов, среди которых наиболее ценен метод вычисления коэффициента корреляции. Эти работы заложили основу для будущего развития генетики человека. Помимо этого Гальтон стал родоначальником евгеники - науки о наследственном здоровье человека и путях его улучшения. Однако принципиальная ошибка Гальтона состояла в том, что в практических мероприятиях евгеники он рекомендовал не столько избавляться от патологических генов, сколько увеличивать количество "хороших" генов в человеческих популяциях путем создания условий для преимущественного размножения одаренных людей. Особого внимания заслуживают исследования известного английского клинициста А. Гэррода (1857 - 1936 гг.), внесшего существенный вклад в изучение проблемы генетики человека. Его работа "Распространенность алкаптонурии: изучение химических особенностей" несла ряд новых идей. Гэррод первым обнаружил взаимосвязь между генами и ферментами, открыл врожденные нарушения обмена веществ и положил начало биохимической генетике. В настоящее время изучение наследственных болезней обмена веществ - наиболее актуальный раздел генетики человека. Труды Гэррода, Адамса и других врачей - исследователей не были оценены при их жизни. Биологи обращали мало внимания на работы медиков. Изучение наследственности проводилось главным образом на растениях. К сожалению, Г. Менделю, как и другим ученым, работавшим с растительными объектами, не были известны данные по генетике человека. В противном случае открытие законов генетики могло бы произойти значительно раньше. В 1865 г. увидела свет знаменитая работа чешского ученого Г. Менделя "Опыты над растительными гибридами". Законы, открытые им, оставались незамеченными в течение 35 лет и только в 1900 г. были переоткрыты К. Корренсом (Германия), Э. Чермаком (Австрия) и Г. де Фризом (Голландия). С тех пор закономерности наследования, открытые Менделем, определяют развитие современной генетики, включая и генетику человека.
Изучая наследования признаков у гороха, Г. Мендель установил три закона:
1. Закон единообразия гибридов первого поколения;
2. Закон расщепления во втором поколении по фенотипу 3:1 (при моногибридном скрещивании);
3.Закон независимого наследования признаков (Шевченко В.А. и др., 2004).
Успех чешского ученого был связан с разработкой принципиально нового методического подхода. Он:
- ввел в науку новый гибридологический метод, выбрав для изучения контрастные пары признаков;
- проводил строгий количественный учет изучаемых признаков, что позволило обнаружить статистические закономерности наследования;
- анализируя эти закономерности, пришел к выводу, что зародышевые клетки несут набор признаков, которые могут быть определены с помощью скрещиваний.
Опыты Г. Менделя и сделанные из них выводы стали предпосылкой для создания теории гена - основы современной генетики, а 1900 г. - год вторичного открытия законов Менделя - считается годом рождения генетики. Название новой науке было дано в 1906 г. английским ученым В.Бэтсоном (от латинского слова geneo - порождаю), а в 1909 г. датский генетик В. Иоганнсен предложил такие важные генетические термины, как ген, генотип и фенотип. В 1903 г. американский антрополог Фараби, изучая родословные в нескольких поколениях, впервые установил, что брахидактилия (короткопалость) у челoвека наследуется по аутосомно-доминантному типу. Из этой работы следовал вывод о справедливости менделевских законов и для человека. В 1900 г. К. Ландштейнер описал систему групп крови АВО. В 1924 г. Ф. Бернштейн установил, что АВО-система групп крови контролируется серией множественных аллелей одного локуса. Спустя 25-30 лет был обнаружен резус-фактор (Rh) и показано, что гемолитическая желтуха новорожденных возникает из-за иммунологической несовместимости матери и плода. Эти открытия также указывали на применимость законов Менделя к наследованию признаков у человека. В 1908 г. Г. Харди и В. Вайнберг независимо друг от друга пришли к выводу, что менделевские законы дают возможность объяснить распределение частоты генов из поколения в поколение в популяциях (от латинского - populus - население, народ) и условиях генетической стабильности популяции. Этот закон был установлен путем анализа наследственности человека и лег в основу популяционной генетики. В 1919 г. Ю.А. Филипченко организовал кафедру генетики в Петроградском университете. В это же время Н.И. Вавилов сформулировал важнейший генетический закон - закон гомологических рядов в наследственной изменчивости. Одновременно в Москве Н.К. Кольцов создает свою генетическую школу. В 20 гг. XX века начала интенсивно развиваться советская генетика. Под влиянием идей евгеники, которая получила широкое распространение в ряде стран Европы (Англия, Франция, Германия) и Америке в 1921 г. в Москве Н.К. Кольцовым было организовано Русское евгеническое общество, в 1922 г. в Петрограде Ю. А. Филипченко создал Бюро по евгенике. Эти евгенические организации ориентировались на сугубо научные задачи в отличие от евгенических обществ других стран. Н.К. Кольцов, Ю.А. Филипченко и другие ученые проводили работы по генетике одаренности, изучая родословные выдающихся личностей. Однако эти исследования грешили методическими ошибками, противоречиями, определенным примитивизмом. Вместе с тем были в евгенических работах и положительные моменты. Так, Н.К. Кольцов и Ю.А. Филипченко правильно ставили вопрос о значении социальных условий в реализации индивидуальных особенностей человека, полностью отвергали насильственный путь улучшения наследственности человека. Кроме того, силами советских евгеников были собраны родословные выдающихся личностей, например, А.С. Пушкина, Л.Н. Толстого, А.М. Горького, Ф.И. Шаляпина и др. К концу 20-х годов евгенические исследования в нашей стране были прекращены. Падала ее популярность и в других странах (кроме Германии). Число евгеничеких обществ быстро уменьшалось, журналы закрывались или переименовывались. Конец 20-х - начало 30-х гг. ознаменовались значительными успехами в развитии генетики. Родилась и стала общепризнанной хромосомная теория наследственности, было установлено, что наследственность связана с генами, локализованными в хромосомах клеточных ядер, что гены в хромосомах расположены линейно и образуют группы сцепления. В этот же период создается популяционная генетика. Большой вклад в развитие этого раздела внесли С.С. Четвериков, Р. Фишер, Н.П. Дубинин и Д.Д. Ромашев, Дж. Е. Холдейн и др. В ряде стран, в том числе в нашей, начинает развиваться медицинская генетика. С 1932 по 37 гг. работал Московский медико-биологический институт им. М. Горького (позднее - Медико-генетический институт), возглавляемый С. Г. Левитом. При нем был организован Центр близнецовых исследований. Здесь изучались болезни с наследственным предрасположением - диабет, язвенная болезнь, аллергия, гипертоническая болезнь и др. Большой интерес имели цитогенетические работы по идентификации первых хромосом человека. Особого упоминания заслуживают труды талантливого генетика и клинициста-невропатолога С.Н. Давиденкова (1880-1961). Он первым поставил вопрос о гетерогенности наследственных заболеваний и начал проводить медико-генетическое консультирование. К концу 30-х гг. XX в. интерес к генетике человека начал снижаться. Сократилось и оставалось низким до начала 50-х гг. количество опубликованных работ. В Советском Союзе с приходом к власти в биологической науке Т.Д. Лысенко все генетические исследования, включая и исследования по генетике человека, были запрещены. Генетика была объявлена "лженаукой". Августовская сессия ВАСХНИЛ (1948 г.) нанесла огромный вред теоретическим и практическим достижениям генетики, утвердив антинаучные идеи Т.Д.Лысенко. Такое положение сохранялось до начала 60-х гг (Шевченко В.А. и др., 2004).
Возрождение советской генетической науки началось после разоблачения "учения" Лысенко и шло по пути развития медицинской генетики. В 1964 г. был издан учебник В.П. Эфроимсона по медицинской генетике, в 1969 г. открыт Институт медицинской генетики под руководством Н.П. Бочкова (в настоящее время - Научно-исследовательский центр медицинской генетики РАМН), где начались широкие исследования по многим направлениям медицинской генетики. В 50-х гг. получают широкое развитие исследования по радиационной генетике человека. Еще в 1927 г. американский исследователь Г. Меллер установил сильное мутагенное действие рентгеновских лучей. Это открытие показало опасность облучения половых клеток человека для последующих поколений, в силу чего человеку как объекту генетических исследований стало уделяться больше внимания. С 1959 по 1962 гг. количество публикаций, симпозиумов, конференций по генетике человека быстро возрастало. Слияние генетики, цитологии, цитогенетики, биохимии способствовало формированию клинической генетики. Усилиями ученых была подтверждена гетерогенность наследственных патологий, когда один и тот же фенотип болезни обусловлен изменением разных белков. Трудно переоценить важность этого открытия для диагностики, лечения и медико-генетического консультирования наследственных болезней. В 1944 г. было достоверно установлено, что передача наследственной информации связана с дезоксирибонуклеино-вой кислотой (ДНК). Это открытие явилось мощным фактором, стимулирующим изучение наследственности на молекулярном уровне. А благодаря созданию в 1953 г. Д. Уотсоном и Ф. Криком модели макромолекулярной структуры ДНК, началось углубленное изучение молекулярной, биохимической и иммуногенетики человека. Убедительный пример значения фундаментальных исследований для практического здравоохранения дает история развития цитогенетики. В 1956 г. X. Тио и А. Леван установили, что в клетках человека содержится 46 хромосом, а спустя три года были открыты хромосомные болезни человека. В 1959 г. Дж. Лежен установил цитогенетическую картину возникновения синдрома Дауна (трисомия по 21-й хромосоме.). В это же время несколько ученых идентифицировали на хромосомном уровне синдром Тернера (ХО) и синдром Клайнфельтера (XXY). Одновременно была определена роль Y-хромосомы в определении пола человека. В 1960 г. Р. Мурхед с коллегами разработали метод культивирования лимфоцитов периферической крови для получения метафазных хромосом человека, что позволило обнаруживать мутации хромосом, характерные для определенных наследственных болезней. Другим важным открытием для развития цитогенетики человека явилась разработка методов дифференциальной окраски хромосом. Благодаря ему стала возможна идентификация каждой хромосомы человека, а это резко повысило разрешающую способность цитогенетических методов. Еще одним этапом развития современной генетики человека явилось картирование и локализация генов в хромосомах человека. Достижения цитогенетики, генетики соматических клеток, увеличение числа генетических маркеров способствовали успешному изучению групп сцепления. В настоящее время у человека установлено 23 группы сцепления. Эти данные нашли непосредственное применение в диагностике наследственных заболеваний и медико-генетическом консультировании. Тесная связь современной генетики с химией, физикой, биохимией, физиологией, экологией, фармакологией и другими науками способствовала появлению новых разделов генетики: цитогенетики, радиационной генетики, иммуногенетики, фармакогенетики, экологической генетики . Во второй половине XX в. начала интенсивно развиваться молекулярная генетика и генная инженерия, были разработаны методы искусственного и ферментативного синтеза генов. В 1969 г. индийский ученый Г. Карано впервые осуществил искусственный синтез гена. С помощью генной инженерии получены искусственные гены инсулина, интерферона, соматотропина и др. Эти достижения открывают большие перспективы в диагностике, профилактике и лечении наследственных болезней человека. Возможности молекулярной генетики и развитие современных методов работы с ДНК нашли применение для решения практических задач медицинской генетики. Конец XX в. ознаменован разработкой и началом осуществления грандиозной международной программы "Геном человека". Ее задача - изучение генома человека, включая картирование хромосом и секвенирование их ДНК, определение полной нуклеотидной последовательности генома, состоящего из трех миллиардов пар нуклотидов. В рамках этой программы разрабатываются методы диагностики и лечения наследственных болезней. В настоящее время уже возможна ДНК- диагностика более 100 наследственных дефектов. В недалеком будущем станет реальностью генотерапия наиболее распространенных болезней человека, патогенез которых уже известен (Шевченко В.А. и др., 2004).
Краткие сведения из истории генетики. Генетика (гр. genetikos — относящийся к происхождению) изучает закономерности наследственности и изменчивости, которые относятся к основным свойствам живой материи, всех организмов. Генетика как наука развилась в связи с практическими потребностями. При разведении домашних животных и культурных растений исстари применялась гибридизация, т. е. скрещивание организмов, относящихся к различным видам, породам, сортам или отличающихся друг от друга какими-либо признаками. Сравнивая гибриды с исходными формами, практики давно подметили некоторые особенности наследования признаков. Основные закономерности преемственности свойств и признаков в поколениях были открыты Г. Менделем (1822—1884). О своих исследованиях он сделал сообщение в 1865 г. на заседании Общества любителей естествознания в г. Брно (Чехословакия). Ставшая впоследствии классической работа Менделя «Опыты над растительными гибридами» была опубликована в трудах того же общества в 1866 г., но в свое время не привлекла внимания современников.
Лишь в 1900 г. те же закономерности вновь установили независимо друг от друга Г. де Фриз в Голландии, К. Корренс в Германии и Э. Чермак в Австрии. Вскоре благодаря опытам по гибридизации, проведенным с многочисленными объектами, было показано, что открытые Менделем закономерности свойственны всем организмам, растениям и животным. 1900 г. Можно считать годом второго рождения генетики (Слюсарев А.А. и др., 1987).
На рубеже XIX и XX вв. передачу наследственных свойств еще не связывали с определенными структурами, хотя не вызывало сомнений, что она осуществляется какими-то факторами материальной природы, находящимися в половых клетках. В начале XX в. по предложению датского ученого И. Иогансена дискретным единицам наследственности было дано название генов. Ко времени вторичного открытия законов Менделя наука уже накопила факты, которые способствовали обнаружению материального субстрата наследственности. В 80-х годах XIX в. Ван-Бенден, Т. Бовери и другие исследователи описали сложные процессы, протекающие при образовании гамет (мейоз), и установили, что в зиготе происходит восста- новление диплоидного набора хромосом, состоящего из одинакового числа хромосом обоих родителей.
Наибольшие успехи были достигнуты школой американского генетика Т. Г. Моргана (1866—1945), сформулировавшего хромосомную теорию наследственности (1911). Школа Моргана доказала, что гены находятся в хромосомах и расположены в них в линейном порядке. Успехи этой школы (как и всего последующего изучения явлений наследственности) в известной мере обусловлены введением нового объекта генетических исследований — плодовой мушки дрозофилы. Для нее характерны интенсивное размножение, быстрая смена поколений (до 30 в год), наличие четко обозначенных альтернативных признаков: цвет тела, величина и форма крыльев, цвет глаз и разведения, относительно несложный кариотип (4 пары хромосом). Как выяснилось в дальнейшем, клетки слюнных желез у дрозофилы содержат особые гигантские хромосомы (политенные). Перечисленные особенности сделали дрозофилу излюбленным объектом для исследований ряда важнейших вопросов генетики.

В начале XX в. господствовало представление о стабильности и неизменяемости генов (А. Вейсман, У. Бэтсон). Считалось также, что если изменение генов возможно, то происходит оно независимо от влияния среды, т. е. самопроизвольно (Г. де Фриз).
Успехи развития биохимии позволили более точно охарактеризовать материальный субстрат наследственности.Советский исследователь Н. К. Кольцов (1872—1940) еще в 1928 г. высказал мысль о связи генов с определенным химическим веществом. Он предполагал, что хромосома представляет собой крупную белковую молекулу, отдельные радикалы которой выполняют функцию генов. Н. К. Кольцов считал, что белковые мицеллы способны к самовоспроизведению. Эта теория оказалась ошибочной, но в ней впервые в науке была сделана попытка рассмотреть закономерности наследственности на молекулярном уровне и впервые, выдвинута идея об авторепродукции единиц наследственной информации (матричный принцип синтеза макромолекул). В последние десятиле наследственности и перейти к выяснению их химической природы (Шевченко В.А и др., 2004).
В 40-х годах Г. Бидл и Е. Татум выяснили, что гены обусловливают образование ферментов, которые, направляя определенным образом клеточный метаболизм, влияют на развитие структур и физиологических свойств организмов (один ген — один фермент).
В 1944 г. О. Эвери, К. Мак-Леод и М. Мак-Карти на микроорганизмах установили, что передача наследственной информации связана с нуклеиновой кислотой (ДНК). Важную роль в изучении ДНК сыграли исследования советского биохимика А. Н. Белозерского (1905—1972). Еще в 30-е годы он представил данные о том, что ДНК — обязательный компонент хромосом клеток растений и животных, и изучил нуклеотидный состав ДНК многих видов. Дальнейшие исследования явлений наследственности должны были перейти на молекулярный уровень. К началу 40-х годов были предложены принципиально новые методы, позволившие заложить основу молекулярной генетики: электронная микроскопия, метод меченых атомов, рентгено-структурный анализ и др. Молекулярная биология возникла на стыке генетики, микробиологии, биохимии и физики. Исследования физиков сыграли важнейшую роль; так, в начале 50-х годов в лаборатории, руководимой английским физиком М. Уилкинсон, с помощью рентгеновских лучей и математических расчетов были получены рентгенограммы нити ДНК. Американский биохимик Р. Чаргафф открыл правило комплементарности пуриновых и пиримидиновых оснований. На основе сопоставления и анализов этих данных генетики Дж. Уотсон и Крик предложили модель макромолекулярной структуры ДНК, имеющей вид двойной спирали. Началось углубленное изучение наследственности на молекулярном уровне, что потребовало привлечения новых объектов исследования. Бактерии, низшие грибы и вирусы стали классическими объектами молекулярной генетики. Благодаря интенсивности размножения и быстроте смены поколений микроорганизмы очень удобны для изучения закономерностей наследственности. Клетка бактерии не является частью организма (как у многоклеточных), а представляет собой особь. Понятие признака и биохимического свойства часто совпадают, т. е. путь от гена к признаку более прямой и прослеживается значительно легче. У прокариот имеется одна хромосома, т. е. любая мутация проявляется фенотипически.
Часто используют в экспериментах кишечную палочку (Escherichia coli), которая входит в состав флоры кишок здорового человека. Она имеет длину 2 мкм и диаметр 1 мкм. Кроме бактерий в молекулярной генетике используют вирусы, в том числе и паразитирующие в клетках бактерий (фаги).
Таким образом, в истории генетики можно выделить три этапа: первый — изучение явлений наследственности на организменном уровне, второй — на клеточном, третий — на молекулярном. Естественно, что и в настоящее время изучение свойств наследственности на всех уровнях не потеряло своего значения. На изучении генетических закономерностей основана селекция, т. е. создание новых и улучшение прежних пород домашних животных, сортов культурных растений, а также микроорганизмов, используемых в фармацевтической промышленности, медицине, народном хозяйстве.
В нашей стране первая кафедра генетики была создана при Петроградском университете в 1919 г. Ю. А. Филипченко (1882—1930), им же написан первый отечественный учебник генетики (Гуттман Б. и др., 2004).
Генетика тесно связана с медициной. В настоящее время известно более двух тысяч наследственных болезней и аномалий развития. Они изучаются на молекулярном, клеточном, организменном и популяциокном уровнях. Генетикой получены важные сведения о том, что наследственные болезни в определенных условиях могут не проявляться; в ряде случаев могут быть даны ценные рекомендации по их предупреждению. Ближайшие задачи медицинской генетики — дальнейшее изучение этих болезней, разработка мероприятий по предупреждению пороков развития, наследственных болезней и злокачественных новообразований.
Всем эукариотическим организмам присущи открытые Г.Менделем общие закономерности наследования признаков. Для их изучения необходимо вспомнить основные термины и понятия, используемые в генетике. Главный постулат Менделя, который он доказал в своих известных экспериментах на горохе огородном, состоит в том, что каждый признак определяется парой наследственных задатков, позже получивших название аллельных генов. С развитием хромосомной теории наследственности выяснилось, что аллельные гены находятся в одинаковых локусах гомологичных хромосом и кодируют один и тот же признак. Пара аллельных генов может быть одинакова (АА или aa), тогда говорят, что особь гомозиготна по данному признаку. Если же аллельные гены в паре разные (Aa), то особь по данному признаку гетерозиготна. Совокупность генов данного организма называется генотипом. Правда часто под генотипом понимают одну или несколько пар аллельных генов, которые отвечают за один и тот же признак. Совокупность признаков данного организма называют фенотипом, фенотип формируется в результате взаимодействия генотипа с внешней средой.
Г. Мендель ввел понятия доминантных и рецессивных генов. Аллель, который определяет фенотип гетерозиготы, он назвал доминантным. Например, ген А в гетерозиготе Аа. Другой аллель, не проявляющий себя в гетерозиготном состоянии, назван им рецессивным. В нашем случае это ген а.
Основные закономерности наследования признаков по Менделю (законы единообразия гибридов первого поколения, расщепления на фенотипичес-кие классы гибридов второго поколения и независимого комбинирования генов) реализуются благодаря существованию закона чистоты гамет.

Г. Мендель (1822-1884)

Менделирующие признаки
Суть последнего состоит в том, что пара аллельных генов, определяющая тот или иной признак: а) никогда не смешивается; б) в процессе гаметогенеза расходится в разные гаметы, то есть в каждую из них попадает один ген из аллельной пары. Цитологически это обеспечивается мейозом: аллельные гены лежат в гомологичных хромосомах, которые в анафазе мейоза расходятся к разным полюсам и попадают в разные гаметы. Генетика человека опирается на общие принципы, полученные первоначально в исследованиях на растениях и животных. Как и у них, у человека имеются менделирующие, т.е. наследуемые по законам, установленным Г. Менделем, признаки. Для человека, как и для других эукариот, характерны все типы наследования: аутосомно-доминантный, аутосомно-рецессивный, наследование признаков, сцепленных с половыми хромосомами, и за счет взаимодействия неаллельных генов. Разработал Г.Мендель и основной метод генетики - гибридологический. Он основан на скрещивании особей одного вида, обладающих альтернативными признаками, и количественном анализе полученных фенотипических классов. Естественно, этот метод не может использоваться в генетике человека (Слюсарев А.А. и др., 1987)
Основные понятия и термины современней генетики.
Наследственностью называется свойство организмов повторять в ряде поколений сходные признаки и обеспечивать специфический характер индивидуального развития определенных условиях среды. Благодаря наследственности родители и потомки имеют сходный тип биосинтеза, определяющий сходство в химическом составе тканей, характере обмена веществ, физиологических отправлениях, морфологических признаках и других особенностях. Вследствие этого каждый вид организмов воспроизводит себя из поколения в поколение. Изменчивость — это явление, противоположное наследственности. Она заключается в изменении наследственных задатков, а также в вариабельности их проявлений в процессе развития организмов при взаимодействии с внешней средой.
Наследственность и изменчивость тесно связаны с эволюцией. В процессе филогенеза органического мира эти два противоположных свойства находятся в неразрывном диалектическом единстве. Новые свойства организмов появляются только благодаря изменчивости, но она лишь тогда может играть роль в эволюции, когда появившиеся изменения сохраняются в последующих поколениях, т. е. наследуются.
Передача наследственных свойств осуществляется в процессе размноже- ния. Размножение, в свою очередь обусловлено делением клеток. При половом размножении передача свойств наследственности осуществляется через половые клетки (гаметы) — яйцеклетки и сперматозооны. При размножении спорообразованием единственным носителем наследственных свойств является спора, при вегетативном размножении — соматические клетки (Слюсарев А.А. и др., 1987).
Элементарными дискретными единицаминаследственности являются гены. С химической точки зрения они представляют собой отрезки молекулы ДНК. Каждый ген определяет последовательность аминокислот в одном из белков, что в конечном счете приводит к реализации тех или иных признаков в онтогенезе особи. Под признаками понимаются морфологические, физиологические, биохимические, патологические и иные свойства организмов, по которым одни из них отличаются от других.
При изучении закономерностей наследования обычно скрещивают организмы, отличающиеся друг от друга альтернативными (взаимоисключающими) признаками. Например, можно взять горох (как это сделал Мендель) с семенами желтыми и зелеными, морщинистыми и гладкими, окраской цветков пурпурной и белой и т. д. Примеры альтернативных признаков у человека: положительный и отрицательный резус-фактор, наличие веснушек и их отсутствие, свободная и приросшая мочка уха и т. д.
Взаимоисключающие признаки являются моногенными, т. е. обычно определяются каким-либо одним геном.
Гены, определяющие развитие альтернативных признаков, принято называть аллельными, или аллеломорфными, парами (гр. allelon — друг с другом, morpha — форма), они расположены в одних и тех же локусах гомологичных хромосом.
Если в обеих гомологичных хромосомах находятся одинаковые аллельные (изоаллельные) гены, такой организм называется гомозиготным и дает только один тип гамет. Если же аллельные гены различны, то такой организм носит название гетерозиготного по данному признаку, он образует два типа гамет.

Совокупность всех наследственных факторов получила название генотипа. Термин «генотип» используется и в более узком смысле для обозначения тех генов, наследование которых составляет предмет изучения.

Совокупность всех признаков и свойств организма называется генотипом. Фенотип развивается на генетической основе в результате взаимодействия организма с условиями внешней среды. Поэтому организмы, имеющие одинаковый генотип, могут отличаться друг от друга в зависимости от условий развития и существования. Пределы, в которых в зависимости от условий среды изменяются фенотипические проявления генотипа, называются нормой реакции.

Процесс передачи наследственной информации от одного поколения к другому получил название наследования. М. Е. Лобашев отмечал, что термины «наследственность» и «наследование» не равнозначны и должны быть четко дифференцированы. Наследственность — общее свойство живого, которое одинаково проявляется у всех организмов, обусловливает хранение и репродукцию наследственной информации, обеспечивает преемственность между поколениями. Так, наследственность — есть свойство живой материи, которое заключено в ее материальности, дискретности и целосности.
Наследование — способ передачи наследственной информации, который может изменяться в зависимости от форм размножения. При бесполом размножении наследование осуществляется через вегетативные клетки и споры, чем обеспечивается большое сходство между материнскими и дочерними поколениями. При половом размножении наследование осуществляется через половые клетки. Сходство между родителями и детьми в этом случае меньше, чем в предыдущем, но зато имеет место большая изменчивость, а следовательно, гораздо более богатый материал для отбора и процесса эволюции.
Основные закономерности наследования. Основные закономерности наследования были открыты Менделем. По уровню развития науки своего времени Мендель не мог еще связать наследственные факторы с определенными структурами клетки. Впоследствии было установлено, что гены находятся в хромосомах, поэтому при объяснении закономерностей, полученных Менделем, мы будем исходить из современных представлений на клеточном уровне. Мендель достиг успеха в своих исследованиях благодаря совершенно новому, разработанному им методу, получившему название гибридологического анализа. Основные положения этого метода следующие:
1. Учитывается не весь многообразный комплекс признаков у родителей и гибридов, а анализируется наследование по отдельным альтернативным признакам.
2. Проводится точный количественный учет наследования каждого альтернативного признака в ряду последовательных поколений: прослеживается не только первое поколение от скрещивания, но и характер потомства каждого гибрида в отдельности. Гибридологический метод нашел широкое применение в науке и практике.
Скрещивание, в котором родительские особи анализируются по одной альтернативной паре признаков, называется моногибридным, по двум — ди-гибридным, по многим альтернативным парам — полигибридным. Прежде всего следует ознакомиться со способом наследования на примере моногибридного скрещивания.

Монозиготное скрещивание
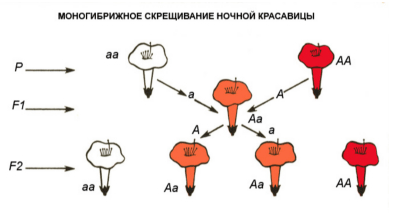
Моногибридное скрещивание. Правило единообразия гибридов первого поколения. В опытах Менделя при скрещивании сортов гороха, имеющих желтые и зеленые семена, все потомство (т. е. гибриды первого поколения) оказалось с желтыми семенами.

Второй закон Менделя

Обнаруженная закономерность получила название правила единообразия гибридов первого поколения. Признак, проявляющийся в первом поколении, получил название доминантного (лат. dominans — господствовать), не проявляющийся, подавленный — рецессивного (лат. recessus — отступление).
«Наследственные факторы» (по современной терминологии — гены) Мендель предложил обозначать буквами латинского алфавита. Гены, относящиеся к одной паре, принято обозначать одной и той же буквой, но доминантный аллель прописной, а рецессивный — строчной. Исходя из сказанного, аллель пурпурной окраски цветов следует обозначать, например, А, аллель белой окраски цветов — а, аллель желтой окраски семян — В, а аллель зеленой окраски семян — b и. т. д.
Вспомним, что каждая клетка тела имеет диплоидный набор хромосом. Все хромосомы парны, аллельные же гены находятся в гомологичных хромосомах. Следовательно, в зиготе всегда имеются два аллеля, и генотипическую формулу по любому признаку необходимо записывать двумя буквами.
Особь, гомозиготную по доминантному аллелю, следует записывать как АА, рецессивную — аа, гетерозиготную — Аа. Опыты показали, что рецессивный аллель проявляется только в гомозиготном состояний, а доминантный — как в гомозиготном, так и в гетерозиготном. Гены расположены в хромосомах. Следовательно, в результате мейоза гомологичные хромосомы (а с ними аллельные гены) расходятся в различные гаметы. Но так как у гомозиготы оба аллеля одинаковы, все гаметы несут один и тот же ген. Таким образом, гомозиготная особь дает один тип гамет.
Опыты по скрещиванию предложено записывать в виде схем. Условились родителей обозначать буквой Р (лат. parentes — родители), особей первого поколения — Fx (лат. filii — дети), особей второго поколения — F2 и т. д. Скрещивание обозначают знаком умножения (X), генотипическую формулу материнской особи записывают первой, а отцовскую — второй. В первой строке выписывают генотипические формулы родителей, во второй — типы их гамет, в третьей — генотипы первого поколения и т. д.
Рассмотрим пример записи при моногибридном скрещивании. Из наблюдений известно, что у человека способность лучше владеть правой рукой доминирует над способностью лучше владеть левой. Если допустим, что в брак вступили гомозиготные правша и левша, то генотипы родителей и детей в этой семье следует записать так:
Р АА X аа
Гаметы А а
F1 Аа 100 %
Поскольку у первого родителя только один тип гамет (А) и у второго родителя также один тип гамет (а), возможно лишь одно сочетание — Аа. Все гибриды первого поколения оказываются однородными: гетерозиготными по генотипу и доминантными по фенотипу.
Следовательно, первый закон Менделя, или закон единообразия гибридов первого поколения, в общем виде можно сформулировать так: при скрещивании гомозиготных особей, отличающихся друг от друга по одной паре альтернативных признаков, все потомство однообразное.
Правило расщепления. При скрещивании однородных гибридов первого поколения между собой (самоопыление или родственное скрещивание) во втором поколении появляются особи как с доминантными, так и с рецессивными признаками, т. е. наблюдается расщепление.
Обобщая фактический материал, Мендель пришел к выводу, что во втором поколении происходит расщепление признаков в определенных частотных соотношениях, а именно: 75 % особей имеют доминантные признаки, а 25 % — рецессивные. Эта закономерность получила название второго правила Менделя, или правила расщепления.

Третий закон Менделя
Согласно второму правилу Менделя, используя современные термины, можно сделать вывод, что: 1) аллельные гены, находясь в гетерозиготном состоянии, не изменяют друг друга; 2) при созревании гамет у гибридов образуется приблизительно равное число гамет с доминантными и рецессивными аллелями; 3) при оплодотворении мужские и женские гаметы, несущие доминантные и рецессивные аллели, свободно комбинируются.
При скрещивании двух гетерозигот (Аа), у каждой из которых образуется два типа гамет — половина с доминантным аллелем (А), половина с рецессивным аллелем (а), следует ожидать четыре возможных сочетания. Яйцеклетка с аллелем А может быть оплодотворена с одинаковой долей вероятности как сперматозооном с аллелем А, так и сперматозооном с аллелем а. Точно так же яйцеклетка с аллелем а может быть оплодотворена сперматозоонами тех же двух типов либо с аллелем А, либо с аллелем а. Получаются зиготы: АА, Аа, Аа, аа. По внешнему облику (фенотипу) особи АА и Аа не отличимы, поэтому расщепление получается в отношении 3:1. Однако по генотипу соотношение остается АА : 2Аа : аа (Шевченко В.А. и др., 2004)

Таким образом, втовое правило Менделя формулируется так: при скрещивании двух гетерозиготных особей, т. е. гибридов, анализируемых по одной альтернативной паре признаков, в потомстве наблюдается расщепление по фенотипу в соотношении 3:1 и по генотипу 1:2:1 Нужно иметь ввиду, что при анализе расщепления в потомстве гибридов фактические числа, полученные из опыта, не всегда соответствуют ожидаемым. Ведь генетические соотношения выражают лишь вероятность появления у потомства определенного признака, а именно вероятность того, что при моногибридном скрещивании во втором поколении должно быть 3/4 особей с доминантными признаками и 1/4 с рецессивными. При малом числе потомков фактические числа могут сильно уклоняться от ожидаемых. Но, как следует из теории вероятности, чем больше фактический материал, тем он точнее выражает истинные соотношения.

Множественный аллелизм
Гипотеза «чистоты гамет». Правило расщепления показывает, что хотя у гетерозигот проявляются лишь доминантные признаки, однако рецессивный ген не утрачен, более того, он не изменился. Следовательно, аллельные гены, находясь в гетерозиготном состоянии, не разбавляются, не изменяют друг друга. Эту закономерность Мендель назвал гипотезой «чистоты гамет». В дальнейшем эта гипотеза получила цитологическое обоснование. Вспомним, что в соматических клетках диплоидный набор хромосом. В одинаковых местах (локусах) гомологичных хромосом находятся аллельные гены. Если это гетерозиготная особь, то в одной из гомологичных хромосом расположен доминантный аллель, в другой — рецессивный. При образовании половых клеток происходит мейоз и в каждую из гамет попадает лишь одна из гомологичных хромосом. В гамете может быть лишь один из аллельных генов. Гаметы остаются «чистыми», они несут только какой-то один из аллелей, определяющий развитие одного из альтернативных признаков.
Доминантные и рецессивные признаки в наследственности человека. В генетике человека известно много как доминантных, так и рецессивных признаков. Одни из них имеют нейтральный характер и обеспечивают полиморфизм в человеческих популяциях, другие приводят к различным патологическим состояниям. Но при этом следует иметь в виду, что доминантные патологические» признаки как у человека, так и у других организмов, если они заметно снижают жизнеспособность, сразу же будут отметены отбором, так как носители их не смогут оставить потомства.
Наоборот, рецессивные гены, даже заметно снижающие жизнеспособность, могут в гетерозиготном состоянии длительно сохраняться, передаваясь из поколения в поколение, и проявляются лишь у гомозигот (Шевченко В.А. и др., 2004).
Анализирующее скрещивание. Генотип организма, имеющего рецессивный признак, определяется по его фенотипу. Такой организм обязательно должен быть гомозиготным по рецессивному гену, так как в случае гетерозиготности у него был бы доминантный признак. Проявляющие доминантные признаки гомозиготная и гетерозиготная особи по фенотипу неотличимы. Для определения генотипа в опытах на растениях и животных производят анализирующие скрещивания и узнают генотип интересующей особи по потомству. Анализирующее скрещивание заключается в том, что особь, генотип которой неясен, но должен быть выяснен, скрещивается с рецессивной формой. Если от такого скрещивания все потомство окажется однородным, значит анализируемая особь гомозиготная, если же произойдет расщепление, то она гетерозиготная.
1. Р АА х аа 2. Р Аа X аа
Гаметы А, а Гаметы А, а а
F1 Аа F1 Аа, аа
Как видно из схемы, при анализирующем скрещивании для потомства гетерозиготной особи характерно расщепление в соотношении 1:1.
Определение генотипов имеет большое значение при селекционной работе в животноводстве и растениеводстве. Анализ генотипов важен также для медицинской генетики. Но в отличие от селекционеров и исследователей, которые имеют дело с растениями и животными и могут ставить эксперименты по скрещиванию организмов, антропогенетик и врач прибегают к анализу родословных и по числовым соотношениям потомков в них которые являются анализирующим.
Поясним примером. У карие глаза доминируют над голубыми следовательно, голубоглазый человек по этому признаку может быть лишь гомозиготным по рецессивным аллелям. Если один из родителей голубоглазый, а другой кареглазый и у них родился голубоглазый ребенок, то следовательно, кареглазый родитель гетерозиготен, если же от родителей с аналогичными признаками родится большое число потомков и у всех будут лишь карие глаза, то нужно полагать, что кареглазый родитель гомозиготен по этому признаку. Второй пример: у обоих родителей полидактилия (многопалость). Ребенок имеет нормальное строение кистей рук. Следовательно, родители гетерозиготны по этому признаку.
Неполное доминирование. В своих опытах Мендель имел дело с призерами полного доминирования, поэтому гетерозиготные особи в его опытах оказались не отличимы от доминантных гомозигот. Но в природе наряду с полным доминированием часто наблюдается неполное, т. е. гетерозиго-ты имеют иной фенотип.
Неполное доминирование
Свойством неполного доминирования обладает ряд генов, вызывающих наследственные аномалии и болезни человека. Например, так наследуются серповидноклеточная анемия (о ней подробнее будет сказано ниже), атаксия Фридрейха. По типу неполного доминирования наследуется цистинурия. У гомозигот по рецессивным аллелям этого гена в почках образуются цистиновые камни, а у гетерозигот обнаруживается лишь повышенное содержание цистина в моче. У гомозигот по гену пильгеровой анемии отсутствует сегментация в ядрах лейкоцитов, а у гетерозигот сегментация есть, но она все же необычная.
Отклонения от ожидаемого расщепления, связанные с летальными генами. В ряде случаев расщепление во втором поколении может отличаться от ожидаемого в связи с тем, что гомозиготы по некоторым генам оказываются нежизнеспособными.
Подобный тип наследования характерен, например, для серых каракульских овец, у которых при скрещивании между собой наблюдается расщепление в соотношении 2:1. Оказалось, что ягнята, гомозиготные по доминантному аллелю серой окраски, гибнут из-за недоразвития пищеварительной системы. У человека аналогично наследуется доминантный ген брахидактилии (укороченные пальцы). Признак проявляется в гетерозиготном состоянии, а у гомозигот этот ген приводит к гибели зародышей на ранних стадиях развития.
Ген серповидно-клеточной анемии кодирует аномальный гемоглобин, проявляющийся и у гетерозигот, но они остаются жизнеспособными, а гомозиготы погибают в раннем детском возрасте. Среди народов Закавказья и Средиземноморья встречается ген талассемии, кодирующий также аномальный гемоглобин. Гомозиготы по нему в 90—95 % случаев погибают, а у гетерозигот отклонение от нормы незначительно.
Концентрация генов аномальных гемоглобинов особенно велика в районах, где прежде была распространена тропическая малярия. Эритроциты с аномальным гемоглобином обладают устойчивостью к проникновению в них малярийных плазмодиев. Носители этих генов малярией не болеют (или болеют в легкой форме). Но в то время, когда не было средств для лечения малярии, они имели преимущество в выживании по сравнению с людьми, имеющими нормальный гемоглобин.
Множественные аллели. Иногда к числу аллельных могут относиться не два, а большее число генов. Они получили название серии множественных аллелей. Возникают множественные аллели в результате многократного мутирования одного и того же локуса в хромосоме. Так, кроме основных доминантного и рецессивного аллелей гена появляются промежуточные аллели, которые по отношению к доминантному ведут себя как рецессивные, а по отношению к рецессивному — как доминантные аллели того же гена.
У кроликов сплошная темная окраска обусловлена доминантным аллелем А, гомозиготные рецессивные животные (аа) — белые. Но существуют еще несколько аллельных состояний этого гена, имеющих собственный фенотип в гомозиготе — шиншилловой (ach, ach) и гималайской (ah, a'1) окраски. Шиншилловые кролики имеют сплошную серую масть. У гималайских кроликов основная масть белая, но кончики ушей, хвоста, ног и носа окрашены.
При скрещивании гималайских кроликов с белыми аллель аь ведет себя по отношению к аллелю а как доминантный. Следовательно, животные с гималайской окраской могут быть двух генотипов: аь аь (гомозиготные) и аь а (гетерозиготные). Но при скрещивании гомозиготного гималайского кролика с шиншилловым аллель аь оказывается рецессивным. Точно так же аллель асЬ проявляет доминантность в отношении не только аллеля аь, но и аллеля а. Следовательно, шиншилловый кролик может быть трех генотипов: ach ach; ach ah; ach а. Аллель А доминирует над всеми другими аллелями этой серии множественных аллелей. Отсюда темной окраске могут соответствовать четыре генотипа: АА, Acfb, Aah, Аа. Вся серия аллелей по фенотипу может быть записана в виде ряда: темный > шиншилла > гималайский > белый или в виде символов: А> ach > ah > а. Таким образом, доминирование — это относительное свойство гена (Слюсарев А.А. и др., 1987).
Наследование одной из групп крови у человека связано с серией множественных аллелей.
Наследование групп крови у человека и явление кодомини-рования. Система групп крови АВО (читается: А, Б, ноль) наследуется по типу множественных аллелей В пределах этой системы имеется четыре фенотипа: группа 1 (0), группа II (Л), группа III (В) и группа IV (АВ). Каждый из этих фенотипов отличается специфическими белками-антигенами, содержащимися в эритроцитах, и антителами — в сыворотке крови. Фенотип I (0) обусловлен отсутствием в эритроцитах антигенов А и В и наличием в сыворотке крови антител а и р. Фенотип II (А) характеризуют эритроциты, содержащие антиген А, и сыворотка крови с антителом р. Фенотип III (В) связан с наличием в эритроцитах антигена В, а в сыворотке крови — антитела а. Фенотип IV {АВ) зависит от наличия в эритроцитах антигенов А и В и отсутствия в сыворотке крови антител а и р.
Установлено, что четыре группы крови человека обусловлены наследованием трех аллелей одного гена (IА , IB, i). При этом нулевая группа обусловлена рецессивным аллелем (і), над которым доминируют как аллель 1А, определяющий II группу, так и аллель /в, от которого зависит III группа. Аллели 1А и 1В в гетерозиготе определяют IV группу, т. е. имеет место кодоминирование. Таким образом, I группа крови бывает лишь при генотипе ii, II — при генотипах 1А1А и IAi, III — при генотипах 1В1В и IBi, IV — при генотипе 1А1В.
Кодоминирование имеет место и при наследовании группы крови по системе MN, открытой в 1927 г. Эта система определяется двумя аллелями: Iм и IN. В сыворотке крови людей с тем или иным фенотипом по этой системе групп крови нет антител к соответствующим антигенам, как это имеет место в системе АВО. Поэтому обычно при переливании крови эта система может не учитываться. Среди европейцев генотип IмIм встречается примерно в 36 %, ININ — в 16 % и 1м1"ъ 48 %.

Наследование групп крови
Принцип наследования групп крови, в частности по системе АВО, используется при спорных случаях в судебной экспертизе с целью исключения отцовства. При этом необходимо помнить следующее. По группам крови нельзя установить, что данный мужчина является отцом ребенка. Можно лишь сказать, мог ли он быть отцом ребенка или отцовство исключено.
Рассмотрим такой признак, как группа крови. Имеется целый ряд типов, или систем, групп крови. Наиболее известна система AB0, по которой различают четыре основных группы: I, II, III и IV; эти группы обозначают также как 0, A, B и AB, поскольку различие между ними определяется тем, какой белок (антиген) присутствует в эритроцитах человека: A или B. Генетически система групп крови AB0 контролируется тремя аллелями: один аллель, обозначаемый A, контролирует синтез антигена A, другой аллель, B, – синтез антигена B, а третий аллель 0 – неактивный и не вызывает образования антигена. Соответственно синтезируемым антигенам и различают четыре группы крови, но им отвечают шесть генетических вариантов (генотипов):
|
генотип |
00 |
A0 AA |
B0 BB |
AB |
|
типы белка |
0 |
A |
B |
AB |
|
группа крови |
I |
II |
III |
IV |
Аллель 0 проявляется фенотипически, т.е. как признак организма, только тогда, когда он оказывается в гомозиготном состоянии (00); этому соответствует первая группа крови, характеризующаяся отсутствием групповых антигенов. В гетерозиготном состоянии (генотипы A0 и B0) он никак не влияет на формируемый фенотип, который целиком определяется альтернативным аллелем (A или B). Поэтому фенотипически генотипы A0 и AA тождественны: они характеризуются наличием антигена A и определяют вторую группу крови. Точно так же тождественны генотипы B0 и BB, определяющие третью группу, т.е. наличие антигена B.
В том случае, когда у гетерозиготной особи фенотипически проявляется только один аллель, говорят, что этот аллель доминантный; при этом другой аллель называется рецессивным. Для системы групп крови AB0, аллели A и B доминируют над аллелем 0; последний же рецессивен по отношению к ним. Если оба аллеля проявляются в фенотипе гетерозиготной особи, то говорят, что они кодоминантны. Так, аллели A и B кодоминантны по отношению друг к другу: в гетерозиготном состоянии (AB) они определяют присутствие обоих антигенов, A и B, т.е. четвертую группу крови.
Механизмы рецессивности и доминантности. Рецессивными часто бывают «дефектные» аллели, не способные производить соответствующий продукт (белок). Поэтому многие наследственные заболевания, обусловленные нехваткой или отсутствием какого-либо белка или фермента, передаются как рецессивный признак: ими страдают только лица гомозиготные по дефектному аллелю. Доминантные болезни чаще всего вызываются аллелями, кодирующими измененные полипептидные цепи. Последние, входя в состав белка, нарушают его пространственную структуру и функциональную активность. Доминантным заболеваниям подвержены лица гетерозиготные по дефектному аллелю. В гомозиготном состоянии доминантные аллели, как правило, летальны.
Расщепление признака в потомстве гетерозигот. У индивидов, гомозиготных по данному гену, все гаметы несут один и тот же его аллель. Среди гамет, производимых гетерозиготной особью, половина несет один аллель, а половина – другой. Знак «плюс» в следующей таблице показывает, какие гаметы производятся разными индивидами по локусу системы групп крови AB0.
|
Группа крови |
I |
II |
III |
IV |
|||
|
Генотип индивида |
00 |
A0 |
AA |
B0 |
BB |
AB |
|
|
Продуцируе- мые гаметы |
0 A B |
+ |
+ + |
+ |
+
+ |
+ |
+ + |
Из этой таблицы видно, что люди со второй и третьей группами крови продуцируют разные гаметы в зависимости от того, гомозиготны они или гетерозиготны. Из таблицы также видно, какой генотип ожидается у детей от родителей с теми или иными группами крови. Если оба родители гомозиготны, то все их дети будут одинаковой группы. Например, родители с первой группой крови образуют гаметы, несущие только аллель 0, поэтому у их детей может быть только первая группа. Если мать имеет вторую, а отец третью группу крови и при этом они гомозиготны, т.е. их генотип, соответственно, АА и ВВ, то дети могут иметь только четвертую группу крови (АВ).
Если же один или оба родителя гетерозиготны, то наблюдается т.н. расщепление признака в потомстве, вытекающее из сформулированного выше Первого правила Менделя и проявляющееся в том, что у детей могут появиться признаки, отсутствующие у родителей. Так, если бы в вышеприведенном примере мать была гетерозиготна, то она производила бы яйцеклетки двух типов – с аллелем A и с аллелем 0. При этом у нее может равновероятно родиться ребенок с третьей либо с четвертой группой крови (генотипа B0 или AB, соответственно). Таким образом, при генотипе матери А0 и отца ВВ дети не могут иметь группу крови матери; их группа крови будет либо такой же, как у отца, либо такой, какая не свойственна ни отцу, ни матери.
Если оба родителя гетерозиготны, то разнообразие генотипов среди детей еще выше. Например, если отец и мать имеют вторую группу крови и генотип их A0, то генотип и группа крови их ребенка зависит от того, какая именно яйцеклетка созрела и каким сперматозоидом она будет оплодотворена. Поскольку в данном примере каждый из родителей производит гаметы A и 0, то генотип их ребенка может быть AA, A0 или 00, а согласно теории вероятностей шансы получить их распределяются как 1:2:1. Поскольку первые два генотипа определяют одну и ту же группу крови, то по признаку «группа крови» шансы иметь ребенка с первой или второй группой крови будут 1:3 (эти соотношения в потомстве гетерозиготных родителей были открыты Менделем). И наконец, если бы мать имела вторую, а отец третью группу крови и оба они были бы гетерозиготными, то с равной вероятностью у них мог бы родиться ребенок с любой группой крови (Шевченко В.А. и др., 2004).
Наследование резус-фактора



Полигибридное скрещивание. Дигибридное скрещивание как пример полигибридного скрещивания.
При полигибридном скрещивании родительские организмы анализируются по нескольким признакам. Примером полигибридного скрещивания может служить дигибридное, при котором у родительских организмов принимаются во внимание отличия по двум парам признаков. Первое поколение гибридов в этом случае оказывается однородным, проявляются только доминантные признаки, причем докинирование не зависит от того, как признаки были распределены между родителями. Вот схема записи двух примеров дигибридного скрещивания:
1. Р ААВВ х aabb
Гаметы АВ ab
F1 AaBb
2. Р ааВВ х ААЪЪ
Гаметы аВ АЬ
F2 AaBb
Правило независимого комбинирования признаков.
При скрещивании гомозиготных особей, отличающихся двумя (или более) парами альтернативных признаков, во втором поколении (F2) наблюдается независимое наследование и комбинирование признаков, если гены, определяющие их, расположены в различных гомологичных хромосомах. Это возможно, так как при мейозе распределение (комбинирование) хромосом в половых клетках при их созревании идет независимо, что может привести к появлению потомков, несущих признаки в сочетаниях, не свойственных родительским и прародительским особям. Рассмотрим это на примере наследования признаков у человека. Предположим, вступают в брак дигетерозиготы по окраске глаз и способности лучше владеть правой рукой (AaBb). При формировании гамет аллель А может оказаться в одной гамете как с аллелем В, так и с аллелем b. Точно так же аллель а может попасть в одну гамету либо с аллелем В, либо с аллелем b. Следовательно, у дигетерозиготной особи образуются четыре возможные комбинации генов в гаметах: АВ, АЬ, аВ, ab. Всех типов гамет будет поровну (по 25%).
Это несложно объяснить поведением хромосом при мейозе. Негомологичные хромосомы при мейозе могут комбинироваться в любых сочетаниях, поэтому хромосома, несущая аллель А, равновероятно может отойти в гамету как с хромосомой, несущей аллель В так и с хромосомой, несущей аллель Ь. Точно так же хромосома, несущая аллель а, может комбинироваться как с хромосомой, несущей аллель В, так и с хромосомой, несущей аллель Ь. Итак, дигетерозиготная особь образует 4 типа гамет. Естественно, что при скрещивании этих гетерозиготных особей любая из четырех типов гамет одного родителя может быть оплодотворена любой из четырех типов гамет, сформированных другим родителем, т. е. возможны 16 комбинаций. Такое же число комбинаций следует ожидать по законам комбинаторики.
При подсчете фенотипов, записанных на решетке Пеннета, оказывается, что из 16 возможных комбинаций во втором поколении в 9 реализуются два доминантных признака (АВ, в нашем примере — кареглазые правши), в 3 — первый признак доминантный, второй рецессивный (Аь, в нашем примере — кареглазые левши), еще в 3 — первый признак рецессивный, второй — доминантный (аВ, т. е. голубоглазые правши), а в одной — оба признака рецессивные (ab, в данном случае — голубоглазый левша). Произошло расщепление по фенотипу в соотношении 9:3:3:1.
Если при дигибридном скрещивании во втором поколении последовательно провести подсчет полученных особей по каждому признаку в отдельности,то результат получится такой же, как при моногибридном скрещивании, т.е. 3 : 1.
В нашем примере при расщеплении по окраске глаз получается •соотношение: кареглазых 12/16, голубоглазых 4/16, по другому признаку — правшей 12/16, левшей 4/16, т. е. известное соотношение 3:1.
Дигетерозигота образует четыре типа гамет, поэтому при скрещивании с рецессивной гомозиготой наблюдается четыре типа потомков; при. этом расщепление как по фенотипу, так и по генотипу происходит в соотношении 1:1:1:1.
Если при скрещивании прослеживается более двух альтернативных признаков, то число ожидаемых комбинаций увеличивается. При тригибридном скрещивании гетерозиготы образуют по 8 типов гамет, дающих 61 сочетания. Рассмотрим этот случай на конкретном примере брака тригетерозигот по характеру лучшего владения правой рукой, окраске глаз и резус-фактору (AaBbRr). На решетке Пеннета представлены все возможные варианты образования зигот: R, зависит полностью от случайности, так как та же хромосома с геном А в равной степени могла встретиться с хромосомой, несущей ген b или г.
В более общей форме, при любых скрещиваниях, расщепление по фенотипу происходит по формуле (3 + 1)n, где п — число пар признаков, принятых во внимание при скрещивании (Шевченко В.А. и др., 2004).
Взаимодействие генов. Развитие любых признаков у организмов является следствием сложных взаимодействий между генами, точнее — между продуктами их деятельности — белками-ферментами. Доминирование проявляется в тех случаях, когда один аллель гена полностью скрывает присутствие второго.
При подсчете фенотипов, полученных в этом случае, наблюдается расщепление в соотношении 27 : 9 : 9 : 9: :3 : 3 : 3 : 1. Это следствие того, что принятые нами во внимание признаки: способность лучше владеть правой рукой, окраска глаз и резус-фактор контролируются генами, локализованными в разных хромосомах, и вероятность встречи хромосомы, несущей ген А, с хромосомой, несущей ген В или другого аллеля. Однако, по-видимому, чаще всего присутствие рецессивного аллеля как-то сказывается, и обычно приходится встречаться с различной степенью неполного доминирования. Это объясняется тем, что доминантный аллель отвечает за активную форму белка-фермента, а рецессивные аллели часто детерминируют те же белки-ферменты, но со сниженной ферментативной активностью. Это явление и реализуется у гетерозиготных форм в виде неполного доминирования.
Сверхдоминирование заключается в том, что у доминантного аллеля в гетерозиготном состоянии иногда отмечается более сильное проявление, чем в гомозиготном состоянии.
Кодоминирование — проявление в гетерозиготном состоянии признаков, детерминируемых обоими аллелями. Например, каждый из ал-'лельных генов кодирует определенный белок, и у гетерозиготного организма синтезируются они оба. В таких случаях путем биохимического исследования можно установить гетерозиготность без проведения анализирующего скрещивания. Этот метод нашел распространение в медико-генетических консультациях для выявления гетерозиготных носителей генов, обусловливающих болезни обмена. По типу кодоминирования у человека наследуется четвертая группа крови.
Сложные отношения возникают между неаллельными парами генов. Комплементарное действие. Комплементарными (лат. соplementum — средство пополнения) называются взаимодополняющие гены, когда для формирования признака необходимо наличие нескольких неаллельных (обычно доминантных) генов. Этот тип наследования в природе широко распространен.

У душистого горошка окраска венчика цветка обусловлена наличием двух доминантных генов (А и В), в отсутствие одного из них — цветки белые. Поэтому при скрещивании растений с генотипами A Abb и ааВВ, имеющих белые венчики, в первом поколении растения оказываются окрашенными, а во втором поколении расщепление происходит в соотношении 9 окрашенных к 7 неокрашенным.

У мышей серая окраска также обусловлена двумя доминантными генами: из них ген А необходим для образования пигмента, поэтому рецессивные гомозиготы (аа) не имеют пигмента (альбиносы). Ген В обусловливает неравномерное распределение пигмента, а рецессивный его аллель (В) кодирует равномерное распределение пигмента по всему волосу и рецессивные гомозиготы (bb) в присутствии гена А имеют черную окраску. Доминантный аллель (В) кодирует отложение пигмента зонами, у вершины и у основания волоса, и носители его в присутствии гена А имеют серую окраску. При скрещивании дигетерозигот (АаВЬ) обнаруживается расщепление в соотношении 9:3:4. Два доминантных гена в генотипе А—В — дают серую окраску, носители доминантного гена А и рецессивного b — черные, а не имеющие гена образования пигмента (ааВ— и aabb) — белые.
Комплементарное взаимодействие генов у человека можно показать на следующих примерах. Нормальный слух обусловлен двумя доминантными неаллельными генами D и Е, из которых один определяет развитие улитки, а другой—слухового нерва. Доминантные гомозиготы и гетерозиготы по обоим генам имеют нормальный слух, рецессивные гомозиготы по одному из этих генов — глухие.
В клетках млекопитающих для защиты от вирусов вырабатывается специфический белок интерферон. Его образование в клетках человека связано с комплементарным взаимодействием двух неаллельных генов, локализованных в разных хромосомах (один — во второй, второй — в пятой хромосоме).
Гемоглобин взрослого человека содержит четыре полипептидные цепи, каждая из которых кодируется отдельным независимым геном. Следовательно, для синтеза молекулы гемоглобина требуется участие четырех комплементарных генов.
У популярного генетического объекта плодовой мушки Drosophila melanogaster имеется большое число форм, наследственно различающихся по окраске глаз. У мух так называемого дикого типа или типа, распространенного в природе, глаза темно-красные. Существуют формы с ярко-красными глазами. Этот признак рецессивен по отношению к дикому типу. Он наследуется по моногибридной схеме при скрещивании нормальных мух и мух с ярко-красными глазами. Соответствующий ген обозначается: st (scarlet) - рецессив; st+ - доминант.
Существуют также мухи с коричневыми глазами. Это тоже рецессивный признак, наследующийся по моногибридной схеме при скрещивании диких мух и мух с коричневыми глазами. Соответствующий ген обозначается bw (brown) - рецессив; bw+ - доминант.
Если скрестить мух с ярко-красными глазами и мух с коричневыми глазами, то получаются следующие результаты. В Fi все мухи имеют темно-красные глаза (дикий тип), а при скрещивании гибридов первого поколения в F2 появляются четыре класса расщепления: мухи с темно-красными, ярко-красными, коричневыми и белыми глазами в соотношении 9:3:3:1. Для объяснения этого результата обратимся к логике генетического анализа.
Результаты, получившиеся в F1, показывают, что существует некоторый тип взаимодействия. Можно предположить, что это взаимодействие аллелей одного гена при моногибридном скрещивании. Однако в F2 появляются четыре класса в соотношении, характерном для дигибридного скрещивания при полном доминировании по обоим признакам, и среди них формируется самый малочисленный класс - мухи с белыми глазами.
Если справедливо предположение о том, что это скрещивание дигибридное, то генотипы родительских форм записывают:
![]()
где знак «+» соответствует нормальным (доминантным) аллелям генов: bw и st. Тогда самки и самцы образуют по одному типугамет - соответственно st bw+ и st+bw, а генотип гибридов F1 будет: st+st bw+bw.
Такие дигетерозиготные мухи должны образовать четыре типа гамет, которые во всевозможных сочетаниях при скрещивании между собой гибридов F1 дадут в F2 следующее расщепление по генотипу.
С помощью фенотипических радикалов можно написать следующее расщепление по фенотипу:
9 st+ - bw+- - с темно-красными глазами
3 st+ - bw bw - с коричневыми глазами
3 st st bw+ - - с ярко-красными глазами
1 st st bw bw - с белыми глазами
Учитывая, что аллели st и bw рецессивны, можно объяснить появление первых трех фенотипических классов при расщеплении. При наличии нормальных аллелей st+ и bw+ мухи должны принадлежать к дикому типу по окраске глаз (9 с темно-красными глазами). При гомозиготности только по рецессивной аллели bw bw мухи должны быть с коричневыми глазами (3), так же как при гомозиготности только по st st мухи должны быть с ярко-красными глазами (3). Наконец, остается последний класс - двойной гомозиготный рецессив (1 st st bw bw), который соответствует мухам с белыми глазами. Все эти выводы можно проверить, исследуя далее расщепление при анализирующих скрещиваниях и скрещиваниях между особями F1.
Таким образом, предположение о дигибридном расщеплении в рассмотренном скрещивании подтверждается, а новообразование - белоглазые мухи в F2 - результат взаимодействия рецессивных аллелей st и bw.
В рассмотренной схеме также наблюдалось взаимодействие генов в F2, в результате которого дрозофилы имели нормальный цвет глаз. Такой тип взаимодействия носит название комплементарности или комплементарного (взаимно дополнительного) действия, когда доминантные аллели обоих генов обусловили нормальный (или дикий) фенотип (под комплементарностью обычно подразумевают именно этот тип взаимодействия генов).
В F2, рецессивные аллели тех же генов обусловили появление белоглазых мух. Был приведен пример того, что носит название формально-генетического анализа, при котором полностью абстрагируются от механизмов действия исследуемых генов. Если анализ проведен верно и формальные отношения генов и аллелей выявлены правильно, то последующее выяснение физиологических механизмов, лежащих в основе генных взаимодействий, только подтверждает и конкретизирует выводы.
Биохимический механизм взаимодействия аллелей генов st и bw исследован достаточно подробно. Известно, что у дрозофилы окраска глаз обусловлена синтезом двух пигментов - красного и бурого. Рецессивная аллель bw в гомозиготе прерывает синтез красного пигмента, поэтому глаза содержат только бурый пигмент. Рецессивная аллель st в гомозиготе блокирует синтез бурого пигмента, вследствие чего в глазах мух содержится только красный пигмент.
Когда в дигетерозиготе оказываются нормальные аллели обоих генов, синтезируются оба пигмента. Результат - комплементарное взаимодействие нормальных аллелей, наблюдаемое в F1. Если в F2 в гомозиготе оказываются и bwbw, и stst, то не синтезируются ни красный, ни бурый пигменты, и глаза оказываются белыми, неокрашенными.

Комплементарное взаимодействие генов, определяющих форму гребня у кур (соотношение 9:3:3:1)
Можно рассмотреть и другой тип комплементарного взаимодействия генов у дрозофилы, если идти от метаболического эффекта генов к схеме их взаимодействия. Вновь обратимся к биосинтезу пигментов глаза у дорозофилы. Известно, что кроме рецессивных аллелей st синтез бурого пигмента блокируют и рецессивные аллели гена purple (pr). Фенотип гомозигот stst и ргрт - ярко-красные глаза. При скрещивании таких мух в F1 глаза нормальные - темно-красные, поскольку работают оба гена - комплементарно взаимодействуют их доминантные аллели. В F2 наблюдается следующее соотношение фенотипов: 9 с темно-красными и 7 с ярко-красными глазами. Это объясняется тем, что выход в гомозиготу любой из двух рецессивных аллелей достаточен для блокирования синтеза пигмента, тем более, когда и рг, и st находятся в гомозиготе. Это тоже пример комплементарного взаимодействия, но без новообразования в F2.
По комплементарному типу взаимодействуют гены, контролирующие разные этапы одного и того же метаболического пути. Однако для многих морфологических признаков неизвестен биохимический механизм их реализации, поэтому приходится ограничиваться констатацией формально-генетической схемы их наследования. Так, по типу комплементарности взаимодействуют гены, определяющие форму гребня кур, форму плода у и др.
Комплементарное взаимодействие генов, определяющих форму плода тыквы (соотношение 9:6: 1)

Эпистаз. Взаимодействие генов, противоположное комплементарному, получило название эпистаза. Под эпистазом понимают подавление неаллельным геном действия другого гена, названного гипостатическим. Например, у кур доминантный аллель гена С обусловливает развитие пигмента, но доминантный аллель другого гена / является его супрессором, «подавителем». Поэтому куры, даже имеющие доминантный аллель гена окраски, в присутствии супрессора оказываются белыми. Следовательно, особи, имеющие в генотипе гены С — белые, а с генотипом СС и Сс — окрашенные. Таким образом, белая окраска кур может быть обусловлена как отсутствием доминантного аллеля гена пигментации, так и наличием доминантного аллеля гена-подавителя окраски. Следовательно, если скрестить двух белых птиц с генотипами (IICC) и (Нес), все особи первого поколения окажутся тоже белыми (Сс), но в F2 при родственном скрещивании особей первого поколения произойдет расщепление по фенотипу в соотношении 13 : 3. Из 16 птиц 3 будут окрашенными (СС i Сс), так как у них ген-супрессор в рецессиве и есть доминантный ген пигментации. Два класса генотипов 9 + 3 окажутся белыми, так как в них присутствует доминантный аллель гена-супрессора; кроме того, рецессивные дигомозиготы (Нес) тоже белые. Сходный фенотип трех классов привел к расщеплению в соотношении 13 : 3.
Вернемся к анализу взаимодействия генов рr и st у дрозофилы. Соотношение фенотипических классов в F2 можно представить себе и как следствие того, что рецессивная аллель рr в гомозиготе препятствует проявлению доминантной аллели st+. Точно так же рецессивная аллель st в гомозиготе препятствует проявлению доминантной аллели рr+. Действительно, то, что известно о генетическом контроле синтеза бурого глазного пигмента у дрозофилы, вполне соответствует предложенному здесь объяснению. Такой тип взаимодействия носит название эпистатического, или эпистаза, и условно изображается: рr > st+ и st > pr+. В данном случае рецессивная аллель рr эпистатична по отношению к доминантной аллели st+, a st эпистатична по отношению к рr+. Данный случай взаимодействия генов называют также двойным рецессивным эпистазом.
По изменению числа и соотношения классов дигибридного расщепления в F2 рассматривают несколько типов эпистатических взаимодействий: простой рецессивный эпистаз (а > В; а> Ь или b > А; Ь > а), который выражается в расщеплении 9:3:4; простой доминаданый эпистаз (А> В; А> b или В > А; B > а) с расщеплением 12:3:1 и т.д.
Один ген, подавляющий действие другого, называют эпистатическим геном, ингибитором или супрессором. Подавляемый ген носит название гипостатического.
Как уже показано, констатация того или иного типа взаимодействия генов в дигибридном скрещивании условна. Тем не менее, при кажущемся нарушении закона независимого наследования (появлении неожиданных классов в расщеплении или уменьшении числа классов), связанного с взаимодействием двух генов, всегда можно свести наблюдаемые соотношения в F2 к классическому 9:3:3:1. При этом важно понять, какие классы объединились, и тогда интерпретировать тип взаимодействия.
Необходимо также отметить, что само словосочетание «взаимдействие генов» условно. В действительности взаимодействуют продукты генов, а не сами гены, так что правильнее было бы говорить о взаимодействии фенов, а не о взаимодействии генов. Отсюда понятно, что судить о том, с каким скрещиванием имеет дело экспериментатор: моногибридным, дигибридным или полигибридным - можно только на основании результатов полного гибридологического анализа.
Полимерия. Различные доминантные неаллельные гены могут оказывать действие на один и тот же признак, усиливая его проявление. Такие гены получили название однозначных, или полимерных, а признаки, ими определяемые,— полигенных. В этом случае два или больше доминантных аллелей в одинаковой степени оказывают влияние на развитие одного и того же признака. Поэтому полимерные гены принято обозначать одной буквой латинского алфавита с указанием цифрового индекса, например: АгАг и axax; А2А2 и а2а2; А3А3 и а3а3 и т. д. Впервые однозначные факторы были обнаружены шведским генетиком Нильсоном-Эле (1908 г.) при изучении наследования окраски у пшениц. Было установлено, что этот признак зависит от двух полимерных генов, поэтому при скрещивании доминантных и рецессивных дигомозигот — окрашенной (А1А1А2А2) с неокрашенной (а1а1а2а2) в F1 все растения дают окрашенные семена, хотя они заметно светлее, чем родительский экземпляр, имеющий красные семена. При скрещивании особей первого поколения обнаруживается расщепление по фенотипу в соотношении 15 : 1, так как бесцветными являются лишь рецессивные дигомозиготы (ахаха2аг). У пигментированных экземпляров интенсивность окраски сильно варьирует в зависимости от числа полученных ими доминантных аллелей: максимальная у доминантных дигомозигот (А1А1А2А2) и минимальная у носителей одного из доминантных аллелей.

Важная особенность полимерии — суммирование (аддитивность) действия неаллельных генов на развитие количественных признаков. Если при моно-генном наследовании признака возможно три варианта «доз» гена в генотипе: АА, Аа, аа, то при полигениом количество их возрастает до четырех и более. Суммирование «доз» полимерных генов обеспечивает существование непрерывных рядов количественных изменений.

Полимерия
Биологическое значение полимерии заключается еще и в том, что определяемые этими генами признаки более стабильны, чем кодируемые одним геном. Организм без полимерных генов был бы крайне неустойчив: любая мутация или рекомбинация приводила бы к резкой изменчиво-.сти, а это в большинстве случаев невыгодно.
У животных и растений имеется много полигенных признаков, среди них и хозяйственно ценные: интенсивность роста, скороспелость, у кур— яйценоскость, у крупного рогатого скота — количество молока, в плодах — содержание сахаристых веществ и витаминов и т. п.
Пигментация кожи у человека определяется пятью или шестью полимерными генами. У коренных жителей Африки (негроидной расы) преобладают доминантные аллели, у представителей европеоидной расы — рецессивные. Поэтому мулаты имеют промежуточную пигментацию, но при вступлении в брак мулатов у них возможно рождение как более, так и менее интенсивно пигментированных детей.

Многие морфологические, физиологические и патологические особенности человека определяются полимерными генами: рост, масса тела, величина артериального давления. Развитие таких признаков у человека подчиняется общим законам полигенного наследования и очень сильно зависит от влияния условий среды. В этих случаях наблюдается, например, наличие предрасположенности к гипертонической болезни, к ожирению и т. п. Данные признаки при благоприятных условиях среды могут и не проявиться или проявиться в незначительной степени. Это отличает поли-генно-наследуемые признаки от моногенных, примеры которых приведены. Изменяя условия среды, можно обеспечить в значительной степени профилактику ряда полигенных заболеваний Комплементарное взаимодействие генов, определяющих форму плода тыквы (соотношение 9:6: 1) (Слюсарев А.А. и др., 1987).
В большинстве случаев отдельные гены, по-видимому, самостоятельно не определяют признаков. В явлениях комплементарности, эпистаза и полимерии обнаруживается фенотипическое выражение молекулярных взаимодействий генов. В ряде экспериментов, проведенных в лабораторных условиях с ферментами, выделенными из организмов с различным генотипом, показано, что механизм комплементарного взаимодействия генов заключается во взаимодействии генных продуктов в цитоплазме.
Наряду с комплементарным и эпистатическим принято также рассматривать взаимодействие генов по типу полимерии. В этом случае разные гены как бы дублируют действие друг друга, и одной доминантной аллели любого из взаимодействующих генов достаточно для проявления изучаемой фенотипической характеристики. Так, при скрещивании растений пастушьей сумки с треугольными плодами (стручками) и с овальными плодами в F1 образуются растения с плодами треугольной формы. При их самоопылении в F2 наблюдается расщепление на растения с треугольными и овальными стручками в соотношении 15:1. Это объясняется тем, что существуют два гена, действующих эднозначно. В этих случаях их обозначают одинаково (А1 и А2). Тогда все генотипы: А1 - А2 -, А1 - а2а2, а2а2А2- будут иметь одинаковую фенотипическую характеристику - треугольные стручки, и только растения а1а1а2а2 будут отличаться - образовывать овальные стручки. Это случай так называемый некумулятивной полимерии.
Однозначные, или полимерные, гены могут действовать и по типу кумулятивной полимерии. Так, шведский генетик Г. Нильсон-Эле в 1908 г. описал серию однозначно действующих генов, которые определяют окраску эндосперма зерен пшеницы. При этом интенсивность окраски зерен оказалась пропорциональной числу доминантных аллелей разных генов в тригибридном скрещивании. Наиболее окрашенными были зерна А1А1А2А2А3А3, а зерна а1а1а2а2а3а3 не имели пигмента. Между этими крайними типами при расщеплении в F2 наблюдались промежуточные варианты в соотношении 1:6:15:20:15:6:1.
По типу кумулятивной полимерии наследуются многие количественные признаки, например цвет кожи у человека; молочность, яйценоскость, масса и другие признаки сельскохозяйственных животных; длина колоса у злаков, содержание сахара в корнеплодах сахарной свеклы и др. Изучением наследования таких признаков занимается специальный раздел генетики - генетика количественных признаков, которая важна прежде всего для селекции и разработки проблем микроэволюции.
Плейотропия. Зависимость нескольких признаков от одного гена носит название плейотропии (гр. pleison — полный, tropos — способ), т. е. наблюдается проявление множественных эффектов одного гена. Это явление было впервые обнаружено Менделем, хотя он специально его не исследовал. По его наблюдениям у растений с пурпурными цветками всегда имелась красная окраска в основании черешков листьев, а кожура семян была бурого цвета. Эти три признака определялись действием одного гена. Н. И. Вавилов описал плейотропное действие гена черной окраски колоса у персидской пшеницы, который вызывал одновременно развитие другого признака — опушение колосковых чешуи. Плейотропное действие гена может быть первичным и вторичным. При первичной плейотропии одновременно проявляет свое множественное действие. Например, измененный белок взаимодействует с цитоплазмой различных клеточных систем, или изменяет свойства мембран в скольких органов. Например, при болезни Хартнепа мутация гена приводит к нарушению всасывания аминокислоты триптофана в кишках и его реабсорбции в почечных канальцах. При этом поражаются одновременно мембраны эпителиальных клеток кишок и почечных канальцев и происходят расстройства пищеварительной и выделительной систем. При вторичной плейотропии имеется одно первичное фенотипическое проявление гена, вслед за которым развивается ступенчатый процесс вторичных проявлений, приводящих к множественным эффектам. Так, при серповидно-клеточной анемии у гомозигот наблюдается несколько патологических признаков: анемия, увеличенная селезенка, поражения кожи, сердца, почек и мозга. Поэтому гомозиготы по гену серповидно-клеточной анемии погибают, как правило, в детском возрасте. Все эти фенотипические проявления гена представляют иерархию вторичных проявлений. Первой причиной, непосредственным фенотипическим проявлением дефектного гена является аномальный гемоглобин и эритроциты серповидной формы. Вследствие этого происходят последовательно другие патологические процессы: слипание и разрушение эритроцитов, анемия, дефекты в почках, сердце, мозге. Эти патологические признаки являются вторичными. По-видимому, большее распространение имеет вторичная плейотропия.
Также изменять, модифицировать проявление других генов, в связи с чем введено понятие о генах-модификатоpax. Последние усиливают или ослабляют развитие признаков, кодируемых «основным» геном. Возможно, что каждый ген является одновременно геном основного действия для «своего» признака и модификатором для других признаков. Таким образом, фенотип — результат взаимодействия генов и всего генотипа с внешней средой в онтогенезе особи.
Учитывая данные, изложенные в этой главе, следует заключить, что не бывает однозначного соотношения между генотипом и фенотипом. Справедливость этого положения подчеркивает и тот факт, что один и тот же ген может в конечном итоге действовать на различные признаки организма.
Первый пример такого множественного, или плейотропного, действия гена содержится в работе Менделя, а именно: окраска цветков и окраска семенной кожуры зависели в его опытах от одного наследственного задатка. У высших растений гены, обусловливающие красную (антоциановую) окраску цветков, одновременно контролируют красную окраску стебля. У человека известен доминантный ген, определяющий признак «паучьи пальцы» (арахнодактилия или синдром Марфана). Одновременно он определяет аномалии хрусталика глаза и порок сердца. В Западном Пакистане обнаружены люди - носители гена, определяющего отсутствие потовых желез на отдельных участках тела. Это одновременно определяет и отсутствие некоторых зубов (Шевченко В.А. и др., 2004)
Признак платиновой окраски шерсти у лисиц контролируется доминантным геном, который существует только в гетерозиготе, поскольку обладает рецессивным летальным действием. При скрещивании платиновых лис наблюдали расщепление на платиновых и серебристо-черных в соотношении 2:1. Такое соотношение может получаться, если платиновые лисицы гетерозиготны (Аа), а черные гомозиготны по рецессивной аллели того же гена (аа). При этом не выживают гомозиготы по доминантной аллели (АА). Такое предположение подтверждается результатами скрещивания платиновых и серебристо-черных лис. Как и следует ожидать, при анализирующем скрещивании получается расщепление на платиновых и серебристо-черных в отношении 1:1. По этой же схеме наследуется наличие (аа) и отсутствие (Аа) чешуи у зеркального карпа, серая (Аа) и черная (аа) окраска каракулевых овец и т.д.
Множественное или плейотропное действие генов связывают с тем, на какой стадии онтогенеза проявляются соответствующие аллели. Чем раньше проявится аллель, тем больше эффект плейотропии.
Учитывая плейотропный эффект многих генов, можно предположить, что часто одни гены выступают в роли модификаторов действия других генов.