БИОЭНЕРГЕТИЧЕСКИЕ ПРОЦЕССЫ: БИОЛОГИЧЕСКОЕ ОКИСЛЕНИЕ, ТКАНЕВОЕ ДЫХАНИЕ. МОЛЕКУЛЯРНАЯ ОРГАНИЗАЦИЯ МИТОХОНДРИАЛЬНОЙ ДЫХАТЕЛЬНОЙ ЦЕПИ. ХЕМИОСМОТИЧЕСКАЯ ТЕОРИЯ ОКИСЛИТЕЛЬНОГО ФОСФОРИЛИРОВАНИЯ. ИНГИБИТОРЫ И РАЗОБЩИТЕЛИ ОКИСЛИТЕЛЬНОГО ФОСФОРИЛИРОВАНИЯ
БИОЭНЕРГЕТИКА
С позиций термодинамики метаболизм представляет собой совокупность процессов, в которой реакции, потребляющие энергию из внешней среды (эндэргонические), сопрягаются с энергодающими (экзэргоническими) реакциями, что позволяет живым существам оказывать постоянное сопротивление нарастанию энтропии. Выяснение биохимических механизмов, приводящих к генерации различных форм биологической энергии, является предметом биоэнергетики. Источником энергии служат реакции, в ходе которых соединения, содержащие атомы углерода в высо-ковосстановленном состоянии, подвергаются окислению , а специальные дыхательные переносчики присоединяют протоны и электроны (восстанавливаются) и в таком виде транспортируют атомы водорода к дыхательной цепи.

Роль АТФ в биоэнергетике
I. БИОЛОГИЧЕСКОЕ ОКИСЛЕНИЕ
Процессы катаболизма в клетках животных сопровождаются потреблением кислорода, который необходим для реакций окисления. В результате этих реакций происходит освобождение энергии, которая необходима организмам в процессах жизнедеятельности для осуществления различных видов работы. Небиологические системы могут совершать работу за счёт тепловой энер-
гии, биологические системы функционируют в изотермическом режиме и для осуществления процессов жизнедеятельности используют химическую энергию. Изучением превращений энергии, сопровождающих химические реакции, занимается биоэнергетика, или биохимическая термодинамика.
СВОБОДНАЯ ЭНЕРГИЯ И ЗАКОНЫ ТЕРМОДИНАМИКИ
Живые организмы с точки зрения термодинамики – открытые системы. Между системой и окружающей средой возможен обмен энергии, который происходит в соответствии с законами термодинамики.
1. Законы термодинамики
Первый закон – закон сохранения энергии; его можно сформулировать так: общая энергия системы и окружающей среды – величина постоянная.
Внутри рассматриваемой системы энергия может переходить от одной её части к другой или превращаться из одной формы в другую.
Второй закон гласит, что все физические и химические процессы в системе стремятся к необратимому переходу полезной энергии в хаотическую, неуправляемую форму. Мерой перехода или неупорядоченности системы служит величина, называемая энтропией (S), она достигает максимума, когда система приходит в истинное равновесие с окружающей средой.
2. Свободная энергия
Каждое органическое соединение, поступающее в организм извне или входящее в состав живой материи, обладает определённым запасом внутренней энергии (Е). Часть этой внутренней энергии может быть использована для совершения полезной работы. Такую энергию системы называют свободной энергией (G).
При постоянных температуре и давлении соотношение между изменением свободной энер гии системы (ΔG) и изменением энтропии (ΔS) можно представить следующим уравнением: ΔG = ΔН – TxS, где ΔН – изменение энтальпии (внутренней энергии или теплоты, содержащейся в системе); Т – абсолютная температура. В условиях, при которых протекают биохимические реакции, АН приблизительно равно ΔЕ (изменению внутренней энергии системы в результате реакции). Для биологических систем измерение свободной энергии производят обычно при стандартных условиях, когда рН 7,0, температура
Рис. Общая схема обмена веществ и энергии. 1 – пищеварение; 2 – катаболизм; 3 – анаболизм; 4 – распад структурно-функциональных компонентов клеток; 5 – экзергонические реакции; 6, 7 – эндерго-нические реакции; 8 – выведение из организма.
При стандартных условиях все функции обозначают как ΔG0‘, ΔS0‘ и ΔН0‘. Изменение стандартной свободной энергии (ΔG0‘) можно вычислить, зная константу равновесия (K’eq) химической реакции.
3. Эндергонические и экзергонические реакции
Направление химической реакции определяется значением AG. Если эта величина отрицательна, то реакция протекает самопроизвольно и сопровождается уменьшением свободной энергии. Такие реакции называют экзергоничес-кими. Если при этом абсолютное значение AG велико, то реакция идёт практически до конца, и её можно рассматривать как необратимую.
Если AG положительно, то реакция будет протекать только при поступлении свободной энергии извне; такие реакции называют эн-дергоническими.
Если абсолютное значение AG велико, то система устойчива, и реакция в таком случае практически не осуществляется. При AG, равном нулю, система находится в равновесии (табл.).
Таблица. Соотношение между величинами K’eq и ΔG0‘ и направлением химических реакций
4. Сопряжение экзергонических и эндергонических процессов в организме
В биологических системах термодинамически невыгодные (эндергонические) реакции могут протекать лишь за счёт энергии экзер-гонических реакций. Такие реакции называют энергетически сопряжёнными. Многие из этих реакций происходят при участии аденозинтрифосфата (АТФ), играющего роль сопрягающего фактора.
Рассмотрим подробнее энергетику сопряжённых реакций на примере фосфорилирования глюкозы.
Реакция фосфорилирования глюкозы свободным фосфатом с образованием глюкозо-6-фосфата является эндергонической:
(1) Глюкоза+ Н3РО4 → Глюкозо-6-фосфат + Н2О (ΔG = +13,8 кДж/моль).
Для протекания такой реакции в сторону образования глюкозо-6-фосфата необходимо её сопряжение с другой реакцией, величина свободной энергии которой больше, чем требуется для фосфорилирования глюкозы.
(2) АТФ → АДФ + Н3РО4 (ΔG = -30,5 кДж/моль).
При сопряжении процессов (1) и (2) в реакции, катализируемой гексокиназой (см. раздел 7), фосфорилирование глюкозы легко протекает в физиологических условиях; равновесие реакции
сильно сдвинуто вправо, и она практически необратима:
(3) Глюкоза + АТФ → Глюкозо-6-фосфат + АДФ (ΔG = -16,7 кДж/моль).
ХАРАКТЕРИСТИКА ВЫСОКОЭНЕРГЕТИЧЕСКИХ ФОСФАТОВ. ЦИКЛ АТФ-АДФ
В живых организмах существует целая группа органических фосфатов, гидролиз которых приводит к освобождению большого количества свободной энергии. Такие соединения называют высокоэнергетическими фосфатами (табл.).
Как видно из таблицы, разные фосфорилированные соединения обладают разным запасом свободной энергии. К группе высокоэнергетических фосфатов, помимо АТФ, относят енолфосфаты, ангидриды и фосфогуанидины. Соединения, расположенные в нижней части таблицы, составляют группу низкоэнергетических фосфатов.
Таблица. Свободная энергия гидролиза некоторых органических фосфатов
Центральное место среди этих соединений занимает АТФ (рис.).
Рис. Аденозинтрифосфорная кислота (АТФ).
В молекуле АТФ две высокоэнергетические (макро-эргические) связи β и γ; они обозначены на рисунке знаком ~ (тильда).
АТФ – молекула, богатая энергией, поскольку она содержит две фосфоангидридные связи (β, γ). При гидролизе концевой фосфоангидридной связи АТФ превращается в АДФ и ортофосфат Pi. При этом изменение свободной энергии составляет -7,3 ккал/моль. При условиях, существующих в клетке в норме (рН 7,0, температура
Таким образом, АТФ – главный, непосредственно используемый донор свободной энергии в биологических системах. В клетке молекула АТФ расходуется в течение одной минуты после её образования. У человека количество АТФ, равное массе тела, образуется и разрушается каждые 24 ч.
Использование АТФ как источника энергии возможно только при условии непрерывного синтеза АТФ из АДФ за счёт энергии окисления органических соединений (рис.).
Рис. Цикл АТФ-АДФ.
Цикл АТФ-АДФ – основной механизм обмена энергии в биологических системах, а АТФ – универсальная «энергетическая валюта».
ОКИСЛИТЕЛЬНО-ВОССТАНОВИТЕЛЬНЫЕ РЕАКЦИИ. ОКИСЛИТЕЛЬНО-ВОССТАНОВИТЕЛЬНЫЙ ПОТЕНЦИАЛ
Под окислением понимают отщепление электронов, а под восстановлением – присоединение электронов. Окисление донора электронов всегда сопровождается восстановлением акцептора электронов. Этот принцип окислительно-восстановительных процессов применим и к биохимическим системам. В любой окислитель-но-восстановительной реакции участвует акцеп-тор электронов (окислитель) и донор электронов (восстановитель). Например:
(1) Си + О → Cu2+O2-.
Суммарную реакцию (1) можно условно разделить на 2 полуреакции (2), (3):
(2) Cu – 2e → Cu2+. (3) О + 2е → О2-.
В каждой из них участвует окисленная и восстановленная форма одного соединения; их называют сопряжённой парой, или редокс-парой.
Разные редокс-пары обладают различным сродством к электрону. Те, у которых это сродство меньше, отдают электрон тем, у кого оно больше. Мерой сродства редокс-пары к электрону служит окислительно-восстановительный потенциал, или редокс-потенциал (Е0‘), величина которого непосредственно связана с изменением свободной энергии. Величину Е0‘ выражают в вольтах; чем она меньше (отрицательнее), тем меньше сродство вещества к электронам. Чем больше сродство, тем больше восстановительный потенциал.
Редокс-потенциалы Е0‘ связаны с изменением свободной энергии уравнением Нернста:
AG0‘ = – nF ΔЕ0 ‘
где- число перенесённых в реакции электронов; F – постоянная Фарадея (23 061 ккал В-1моль-1); ΔЕо‘ – разность редокс-потенциалов электрон-донорной и электрон-акцепторной пар.
Величина ΔЕо‘ – стандартная величина окислительно-восстановительного потенциала; её определяют в стандартных условиях, когда концентрации всех веществ равны
ОСНОВНЫЕ ЭТАПЫ ТРАНСФОРМАЦИИ ЭНЕРГИИ КАТАБОЛИЧЕСКИХ ПРОЦЕССОВ
Энергия освобождается в процессе ферментативного окисления метаболитов специфическими дегидрогеназами. В реакциях дегидрирования электроны и протоны переходят от органических субстратов на коферменты NAD- и FAD-зависимых дегидрогеназ. Электроны, обладающие высоким энергетическим потенциалом, передаются от восстановленных коферментов NADH и FADH2 к кислороду через цепь переносчиков, локализованных во внутренней мембране митохондрий. Восстановление молекулы О2 происходит в результате переноса 4 электронов. При каждом присоединении к кислороду 2 электронов, поступающих к нему по цепи переносчиков, из матрикса поглощаются 2 протона, в результате чего образуется молекула Н2О.
Окисление органических веществ в клетках, сопровождающееся потреблением кислорода и синтезом воды, называют тканевым дыханием, а цепь переноса электронов (ЦПЭ) – дыхательной цепью.
Электроны, поступающие в ЦПЭ, по мере их продвижения от одного переносчика к другому теряют свободную энергию. Значительная часть этой энергии запасается в форме АТФ, а часть энергии рассеивается в виде тепла. Кроме того, электроны с высоким энергетическим потенциалом, возникающие при окислении различных субстратов, могут быть использованы в реакциях биосинтеза, для которых помимо АТФ требуются восстановительные эквиваленты, например NADPH.
ФЕРМЕНТЫ И КОФЕРМЕНТЫ, УЧАСТВУЮЩИЕ В ОКИСЛИТЕЛЬНО-ВОССТАНОВИТЕЛЬНЫХ РЕАКЦИЯХ
Перенос электронов от окисляемых субстратов к кислороду происходит в несколько этапов. В нём участвует большое количество промежуточных переносчиков, каждый из которых способен присоединять электроны от предыдущего компонента и передавать следующему. Так возникает цепь окислительно-восстановительных реакций, в результате чего происходят восстановление О2 и синтез Н2О. В дыхательную цепь митохондрий входит большое число переносчиков (рис.).
Рис. Митохондриальная цепь переноса электронов. Комплекс I содержит FMN и не менее пяти железо-серных белков (FeS). Комплекс III включает две разные формы цитохрома b (с максимумами поглощения 562 и 566), один FeS-белок и цитохром с1. Комплекс IV содержит цитохромы а и а3 и два иона меди. Комплекс II (сукцинатдегидрогеназа). Комплекс V – АТФ-синтаза.
За исключением убихинона (КоQ), все компоненты ЦПЭ – белки. В составе этих белков содержатся различные небелковые компоненты: FMN, Fе в составе железо-серных белков и в составе порфириновых колец, ионы Cu.
1. Первичные акцепторы водорода
Первичные акцепторы водорода окислительно-восстановительных реакций относят к 2 типам де-гидрогеназ: никотинамидзависимым, содержащим в качестве коферментов производные никотиновой кислоты, и флавинзависимым, содержащим производные рибофлавина.
Никотинамидзависимые дегидрогеназы содержат в качестве коферментов NAD+ или NADР+ (см. раздел 2). NAD+ и NADР+ – производные витамина РР. Эти коферменты входят в состав активных центров дегидрогеназ, но могут обратимо диссоциировать из комплекса с апофер-ментами и включаются в состав фермента в ходе реакции. Субстраты NAD- и NADР-зависимых дегидрогеназ находятся в матриксе митохондрий и в цитозоле. Рабочей частью никотинамидных коферментов служит никотинамид (рис.).
Рис. Структурные формулы рабочей части коферментов NAD+ и NADP+. В окисленной форме никотинамид-ные коферменты обозначают как NAD+ и NADP+, так как они несут положительный заряд на атоме азота пиридинового кольца. В реакциях дегидрирования из двух атомов водорода, отщепляемых от окисляемого субстрата, никотинамидное кольцо присоединяет ион водорода и два электрона в форме гидрид-иона (:Н–). Второй ион переходит в среду. В обратной реакции NADH (NADPH) выступают в качестве доноров электронов и протонов.
Большинство дегидрогеназ, поставляющих электроны в ЦПЭ, содержат NAD+. Они катализируют реакции типа:
R-CHOH-R1 + NAD+ ↔ R-CO-R1 + NADH + H+.
Таким образом, NAD+, присоединяя протоны и электроны от различных субстратов, служит главным коллектором энергии окисляемых веществ и главным источником электронов, обладающих высоким энергетическим потенциалом, для ЦПЭ.
NADРН не является непосредственным донором электронов в ЦПЭ, а используется почти исключительно в восстановительньгх биосинтезах. Однако возможно включение электронов с NADPH в ЦПЭ благодаря действию пиридиннуклеотид трансгидрогеназы, катализирующей реакцию:
NADPH + NAD+ ↔ NADP+ + NADH.
Флавиновые дегидрогеназы содержат в качестве коферментов FAD или FMN. Эти коферменты образуются в организме человека из витамина В2. Флавиновые коферменты прочно связаны с апоферментами. Рабочей частью FAD и FMN служит изоаллоксазиновая сопряжённая циклическая система (рис.).
Рис. Структурные формулы рабочей части коферментов FAD и FMN. В ходе реакции FAD и FMN присоединяют 2 электрона и, в отличие от NAD+, оба теряемых субстратом протона.
FAD служит акцептором электронов от многих субстратов в реакциях типа:
R-CH2-CH2-R1 + E (FAD) ↔ R-CH=CH-R1 + E (FADH2),
где Е – белковая часть фермента.
Большинство FAD-зависимых дегидрогеназ – растворимые белки, локализованные в матриксе митохондрий. Исключение составляет сукци-натдегидрогеназа, находящаяся во внутренней мембране митохондрий. К FMN-содержащим
ферментам принадлежит NADH-дегидрогена-за, которая также локализована во внутренней мембране митохондрий; она окисляет NADH, образующийся в митохондриальном матриксе.
2. Цепь переноса электронов от NADH и FADH2 на кислород
Перенос электронов от NADH к О2 включает ряд переносчиков, которые локализованы во внутренней мембране митохондрий. За исключением убихинона и цитохрома С, это сложные белковые комплексы.
NADH-дегидрогеназа (NADH-Q-редуктаза, комплекс I) состоит из нескольких полипептидных цепей. Роль простетической группы играет FMN. Единственный субстрат фермента – NADH, с которого 2 электрона и протон переносятся на FMN с образованием FMNH2. Второй протон поглощается из матрикса. Реакция протекает по уравнению:
NADH + H+ + Е (FMN) → NAD+ + Е (FMNH2).
С FMNH2 электроны переносятся затем на ряд железо-серных белков (FeS), играющих роль второй простетической группы в молекуле NADН-дегидрогеназы. Атомы железа в этих белках (негемовое железо) собраны в несколько групп, так называемых железо-серных центров. FeS-центры входят в состав многих белков (флавопротеинов, цитохромов), участвующих в окислительно-восстановительных реакциях. Известны 3 типа FeS-центров (FeS, Fe2S2, Fe4S4), в которых атом железа связан с атомом серы остатков цистеина или неорганической серы. Строение железо-серных центров показано на рис.
Рис. Строение железо-серных центров. I – FeS-центр; атом железа связан координационными связями с четырьмя атомами серы, принадлежащими четырём остаткам цистеина в белке. II – Fe2S2-центр; каждый из двух атомов железа связан координационными связями с двумя атомами неорганической серы и двумя остатками цистеина в белке; III – Fe4S4-центр; четыре атома железа связаны с четырьмя атомами серы и четырьмя остатками цистеина в белке. Атомы железа в FeS-центрах могут находиться в окисленном (Fe3+) или восстановленном (Fe2+) состоянии.
NADН-дегидрогеназа содержит несколько центров типа Fe2S2 и Fe4S4. Атомы железа в таких центрах могут принимать и отдавать электроны поочерёдно, переходя в ферро- (Fe2+) и ферри-(Fe3+) состояния. От железо-серных центров электроны переносятся на кофермент Q (убихинон) (рис.).
Рис. Структура убихинона (кофермента Q). n – число изопреноидных звеньев. Убихинон может принимать один электрон и превращаться в семихинон или 2 электрона и полностью восстанавливаться в гидрохинон (убихинол).
Обозначение этого жирорастворимого хинона происходит от первой буквы английского названия хинона (quinone), а название убихинон отражает его широкую распространённость в природе (ubiquitous – вездесущий). Молекулы убихинона в зависимости от источника, из которого они выделены, различаются длиной углеводородной цепи, которая у млекопитающих содержит 10 изопреноидных звеньев и обозначается как Q10. В процессе переноса электронов с NADН-дегидрогеназы через FeS на убихинон он обратимо превращается в гидрохинон. Убихинон выполняет коллекторную функцию, присоединяя электроны от NADН-дегидрогеназы и других флавинзависимых дегидрогеназ, в частности, от сукцинатдегидрогеназы. Убихинон участвует в реакциях типа:
Е (FMNH2) + Q → E (FMN) + QH2.
Цитохромы или гемопротеины присутствуют во всех типах организмов. В клетках эукариотов они локализованы в митохондриальных мембранах и в ЭР. Известно около 30 различных цитохромов. Все цитохромы в качестве простетической группы содержат гем (см. раздел 1). Их многообразие обусловлено:
• различием боковых цепей в структуре гема;
• различием в структуре полипептидных цепей;
• различием в способе связи полипептидных цепей с гемом.
В зависимости от способности поглощать свет в определённой части спектра все цитохромы делят на группы а, b, с. Внутри каждой группы отдельные виды с уникальными спектральными свойствами обозначают цифровыми індексами (b, b1, b2 и т.д.).
Структурные особенности разных видов цитохромов определяют различие в их окислительно-восстановительных потенциалах. В ЦПЭ участвуют 5 типов цитохромов (a, a3, b, с, с1).
За исключением цитохрома с, все цитохромы находятся во внутренней мембране митохондрий в виде сложных белковых комплексов (табл.).
Таблица. Компоненты митохондриальной цепи переноса электронов
QН2-дегидрогеназа (коэнзим Q-цитохром с-редуктаза, комплекс III) состоит из 2 типов цитохромов (b1 и b2) и цитохрома с1. QН2-дегидрогеназа переносит электроны от убихинола на цитохром с. Внутри комплекса III электроны передаются от цитохромов b на FeS-центры, на цитохром с1, а затем на цитохром с. Группы гема, подобно FeS-центрам, переносят только по одному электрону. Таким образом, от молекулы 2 электрона переносятся на 2 молекулы цитохромов b. В качестве промежуточного продукта в этих реакциях переноса электронов возможно образование свободного радикала семихинона. В цитохромах типа b гем не связан ковалентно с белком, а в цитохромах с1 и с он присоединяется к белку при помощи тиоэфир-ных связей (рис.).
Рис. Структура гема цитохромов b, с, с1.
Эти связи образуются путём присоединения 2 цистеиновых остатков к винильным группам гема.
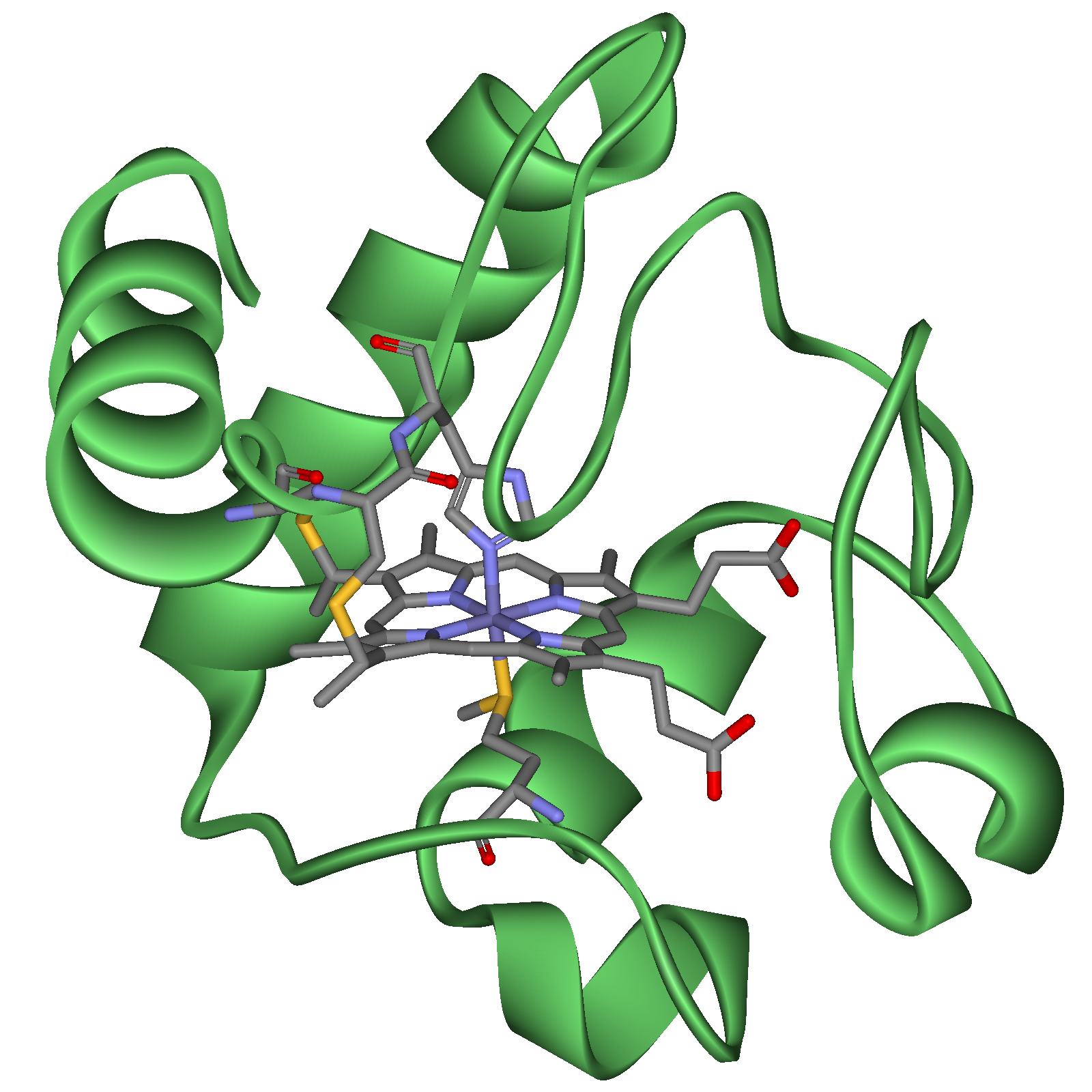
Цитохром с – периферический водорастворимый мембранный белок с молекулярной массой 12 500 Д, имеющий одну полипептидную цепь из 100 аминокислотных остатков, и молекулу гема, ковалентно связанную с полипептидом.
Цитохромоксидаза (комплекс IV) состоит из 2 цитохромов типа аа3, каждый из которых имеет центр связывания с кислородом. Цитохромы а и а3 имеют характерную железопорфириновую простетическую группу, называемую гемом А и отличающуюся от гема цитохромов с и с1 (рис. 6-10). Он содержит формильную группу вместо одной из метильных групп и углеводородную цепь вместо одной из винильных групп.
Другая особенность комплекса а-а3 – наличие в нём ионов меди, связанных с белковой частью в так называемых СиА-центрах. Перенос электронов комплексом a-а3, включает реакции:
Cu+ ↔ Cu2+ + е,
Fe2+ ↔ Fe3+ + e.
Комплекс цитохромов а-а3 непосредственно реагирует с молекулярным кислородом.
ОРГАНИЗАЦИЯ ДЫХАТЕЛЬНОЙ ЦЕПИ В МИТОХОНДРИЯХ
Основные переносчики электронов встроены во внутреннюю мембрану митохондрий и организованы в 4 комплекса, расположенных в определённой последовательности (векторно). В этой последовательности их стандартные окислительно-восстановительные потенциалы становятся более положительными по мере приближения к кислороду Каждое звено этой цепи специфично в отношении донора и акцептора электронов.
На первом этапе дегидрогеназы катализируют отщепление водорода от различных субстратов. Если субстратами служат α-гид-роксикислоты малат, изоцитрат, 3-гидрокси-бутират, водород переносится на NAD+. Образовавшийся NADH в дыхательной цепи, в свою очередь, окисляется NADH-дегидро-геназой (комплекс I).
Если субстратом служат такие соединения, как сукцинат или глицерол-3-фосфат, акцептором водорода служат FAD-зависимые дегидрогеназы. От NADH и FADH2 электроны и протоны передаются на убихинон и далее через цепь цитохромов к молекулярному кислороду.
До сих пор точно неизвестно, каким образом расположены все переносчики электронов дыхательной цепи. Однако установлено, что в расположении дыхательных комплексов существует определённая асимметрия: некоторые из белков-переносчиков находятся ближе к той стороне внутренней мембраны, которая обращена к матриксу, а другие – к противоположной; некоторые белки пронизывают мембрану насквозь.
ОКИСЛИТЕЛЬНОЕ ФОСФОРИЛИРОВАНИЕ АДФ
Так как электроны всегда стремятся переходить от электроотрицательных систем к электроположительным, их транспорт по ЦПЭ к кислороду сопровождается снижением свободной энергии.
Вместе с тем в дыхательной цепи можно выделить 3 участка, в которых перенос электронов сопровождается относительно большим снижением свободной энергии Эти этапы способны обеспечить энергией синтез АТФ, так как количество выделяющейся свободной энергии приблизительно равно энергии, необходимой для синтеза АТФ из AДФ и фосфата. Экспериментально было подтверждено, что процесс переноса электронов по ЦПЭ и синтез АТФ энергетически сопряжены.
Первый процесс – перенос электронов от восстановленных коферментов NADH и FADH2 через ЦПЭ на кислород – экзергонический. Например:
NADH + H+ + 1/2 O2 → NAD+ + H2O + 52 ккал/моль(=220 кДж/моль).
Второй процесс – фосфорилирование AДФ, или синтез АТФ, – эндергонический:
АДФ + Н3РО4+7,3 ккал/моль (30,5 кДж/моль) = АТФ + Н2О.
Синтез АТФ из АДФ и Н3РО4 за счёт энергии переноса электронов по ЦПЭ называют окислительным фосфорилированием.
МЕХАНИЗМ СОПРЯЖЕНИЯ ОКИСЛЕНИЯ И ФОСФОРИЛИРОВАНИЯ
Каким же образом осуществляется сопряжение этих двух процессов? Наиболее обоснованный ответ на этот вопрос даёт хемиосмотическая теория Митчелла, предложенная им в
1. Протонный градиент и электрохимический потенциал
Перенос электронов по дыхательной цепи от NADH к кислороду сопровождается выкачиванием протонов из матрикса митохондрий через внутреннюю мембрану в межмембранное пространство. На эту работу затрачивается часть энергии электронов, переносимых по ЦПЭ.
Протоны, перенесённые из матрикса в межмембранное пространство, не могут вернуться обратно в матрикс, так как внутренняя мембрана непроницаема для протонов. Таким образом, создаётся протонный градиент, при котором концентрация протонов в межмембранном пространстве больше, а рН меньше, чем в матриксе. Кроме того, каждый протон несёт положительный заряд, и вследствие этого появляется разность потенциалов по обе стороны мембраны: отрицательный заряд на внутренней стороне и положительный – на внешней. В совокупности электрический и концентрационный градиенты составляют электрохимический потенциал ΔμН+ – источник энергии для синтеза АТФ. Так как наиболее активный транспорт протонов в межмембранное пространство, необходимый для образования ΔμН+, происходит на участках ЦПЭ, соответствующих расположению комплексов I, III и IV, эти участки называют пунктами сопряжения дыхания и фосфорилирования (рис.).
Рис. Сопряжение дыхания и синтеза АТФ в митохондриях. I – NADH-дегидрогеназа; II – сукцинат дегидрогеназа; III – QH2-дегидрогеназа; IV – цитохромоксидаза; V – ЛТФ-синтаза. Энергия протонного потенциала (электрохимического потенциала ΔμН+) используется для синтеза АТФ, если протоны возвращаются в матрикс через ионные каналы АТФ-синтазы.
Механизм транспорта протонов через митохондриальную мембрану в пунктах сопряжения недостаточно ясен. Однако установлено, что важную роль в этом процессе играет KoQ. Наиболее детально механизм переноса протонов при участии KoQ изучен на уровне комплекса III (рис.).
Рис. Сопряжение переноса электронов через дыхательный комплекс III с транспортом H+ через мембрану. Восстановленный убихинон (QH2) взаимодействует с Fe3+ гема b1 и, восстанавливая его, освобождает протон в водную фазу, превращаясь в семихинон (HQ*). Электрон от гема b1 переносится на Fe3+ гема b2. HQ* отдаёт второй электрон на FeS-центр, расположенный ближе к наружной поверхности мембраны; при этом второй протон оказывается в межмембранном пространстве; электрон передаётся на цитохром с1, а далее на цитохром с. Окисленный Q диффундирует к внутренней стороне мембраны, где получает электрон от гема b2 и протон из матрикса, превращаясь в HQ*. HQ* получает электрон от комплекса I и протон из матрикса; в мембране образуется QH2, и весь процесс повторяется сначала.
KoQ переносит электроны от комплекса I к комплексу III и протоны из матрикса в межмембранное пространство, совершая своеобразные циклические превращения, называемые Q-циклами. Донором электронов для комплекса III служит восстановленный убихинон (QH2), а акцептором – цитохром с. Цитохром с находится с внешней стороны внутренней мембраны митохондрий; там же располагается активный центр цитохрома с1, с которого электроны переносятся на цитохром с.
В мембране существует стационарный общий фонд Q/QH2, из которого каждая молекула QH2 в одном цикле обеспечивает перенос протонов из матрикса в межмембранное пространство и электронов, которые в конечном итоге поступают на кислород. На работу, совершаемую при выкачивании протонов, расходуется часть свободной энергии, которая освобождается при переносе электронов по градиенту редокс-по-тенциала. Энергия электрохимического потенциала (ΔμН+) используется для синтеза АТФ, если протоны возвращаются в матрикс через ионные каналы АТФ-синтазы.
2. Строение АТФ-синтазы и синтез АТФ
АТФ-синтаза (Н+-АТФ-аза) – интегральный белок внутренней мембраны митохондрий. Он расположен в непосредственной близости к дыхательной цепи. АТФ-синтаза состоит из 2 белковых комплексов, обозначаемых как F0 и F1 (рис.).
Рис. Строение и механизм действия АТФ-синтазы. А – F0 и F1 – комплексы АТФ-синтазы. В состав F0 входят полипептидные цепи, которые образуют канал, пронизывающий мембрану насквозь. По этому каналу протоны возвращаются в матрикс из межмембранного пространства; белок F1 выступает в матрикс с внутренней стороны мембраны и содержит 9 субъединиц, 6 из которых образуют 3 пары α и β («головка»), прикрывающие стержневую часть, которая состоит из 3 субъединиц γ, δ и ε. γ и ε подвижны и образуют стержень, вращающийся внутри неподвижной головки и связанный с комплексом F0. В активных центрах, образованных парами субъединиц α и β, происходит связывание AДФ, неорганического фосфата (Р) и АТФ. Б – Каталитический цикл синтеза АТФ включает 3 фазы, каждая из которых проходит поочерёдно в 3 активных центрах: 1 – связывание АДФ и Н3РО4; 2 – образование фосфоангидридной связи АТФ; 3 – освобождение конечного продукта. При каждом переносе протонов через канал F0 в матрикс все 3 активных центра катализируют очередную фазу цикла. Энергия электрохимического потенциала расходуется на поворот стержня, в результате которого циклически изменяется конформация α- и β-субъединиц и происходит синтез АТФ.
Гидрофобный комплекс F0 погружён в мембрану. Он служит основанием, которое фиксирует АТФ-синтазу в мембране. Комплекс F0 состоит из нескольких субъединиц, образующих канал, по которому протоны переносятся в матрикс.
Комплекс F1 выступает в митохондриальный матрикс. Он состоит из 9 субъединиц (3α, 3β, γ, ε, δ). Субъединицы α и β уложены попарно, образуя «головку»; между α- и β-субъединицами располагаются 3 активных центра, в которых происходит синтез АТФ; γ-, ε-, δ- субъединицы связывают комплекс F1 с F0.
Повышение концентрации протонов в межмембранном пространстве активирует АТФ-синтазу. Электрохимический потенциал ΔμН+ заставляет протоны двигаться по каналу АТФ-синтазы в матрикс. Параллельно под действием ΔμН+ происходят конформационные изменения в парах α, β-субъединиц белка F1, в результате чего из AДФ и неорганического фосфата образуется АТФ. Электрохимический потенциал,
генерируемый в каждом из 3 пунктов сопряжения в ЦПЭ, используют для синтеза одной молекулы АТФ.
3. Коэффициент окислительного фосфорилирования
Окисление молекулы NADH в ЦПЭ сопровождается образованием 3 молекул АТФ; электроны от FAD-зависимых дегидрогеназ поступают в ЦПЭ на KoQ, минуя первый пункт сопряжения. Поэтому образуются только 2 молекулы АТФ. Отношение количества фосфорной кислоты (Р), использованной на фосфорилирование AДФ, к атому кислорода (О), поглощённого в процессе дыхания, называют коэффициентом окислительного фосфорилирования и обозначают Р/О. Следовательно, для NADH Р/О = 3, для сукцината Р/О = 2. Эти величины отражают теоретический максимум синтеза АТФ, фактически эта величина меньше.
4. Дыхательный контроль
Окисление субстратов и фосфорилирование AДФ в митохондриях прочно сопряжены. Скорость использования АТФ регулирует скорость потока электронов в ЦПЭ. Если АТФ не используется и его концентрация в клетках возрастает, то прекращается и поток электронов к кислороду.
С другой стороны, расход АТФ и превращение его в AДФ увеличивает окисление субстратов и поглощение кислорода. Зависимость интенсивности дыхания митохондрий от концентрации AДФ называют дыхательным контролем. Механизм дыхательного контроля характеризуется высокой точностью и имеет важное значение, так как в результате его действия скорость синтеза АТФ соответствует потребностям клетки в энергии. Запасов АТФ в клетке не существует. Относительные концентрации АТФ/АДФ в тканях изменяются в узких пределах, в то время как потребление энергии клеткой, т.е. частота оборотов цикла АТФ и AДФ, может меняться в десятки раз.
Общее содержание АТФ в организме 30-
ТРАНСПОРТ АТФ И АДФ ЧЕРЕЗ МЕМБРАНЫ МИТОХОНДРИЙ
В большинстве эукариотических клеток синтез основного количества АТФ происходит внутри митохондрии, а основные потребители АТФ расположены вне её. С другой стороны, в матриксе митохондрий должна поддерживаться достаточная концентрация AДФ. Эти заряженные молекулы не когут самостоятельно пройти через липидный слой мембран. Внутренняя мембрана непроницаема для заряженных и гидрофильных веществ, но в ней содержится определённое количество транспортёров, избирательно переносящих подобные молекулы из цитозоля в матрикс и из матрикса в цитозоль.
В мембране есть белок АТФ/AДФ-антипортер, осуществляющий перенос этих метаболитов через мембрану (рис.).
Рис. Схема трансмембранного переноса веществ за счёт энергии АцН. Потоки различных веществ (АТФ, AДФ, Н3РО4, Са2+) проходят через специфические транспортёры, при этом затрачивается энергия электрохимического потенциала мембраны.
Молекула АДФ поступает в митохондриальный матрикс только при условии выхода молекулы АТФ из матрикса.
Движущая сила такого обмена – мембранный потенциал переноса электронов по ЦПЭ. Расчёты показывают, что на транспорт АТФ и AДФ расходуется около четверти свободной энергии протонного потенциала. Другие транспортёры тоже могут использовать энергию электрохимического градиента. Так переносится внутрь митохондрии неорганический фосфат, необходимый для синтеза АТФ. Непосредственным источником свободной энергии для транспорта Са2+ в матрикс также служит протонный потенциал, а не энергия АТФ.
Биологические виды энергии. Энергетические превращения в живой клетке подразделяют на две группы: локализованные в мембранах и протекающие в цитоплазме. В каждом случае для «оплаты» энергетических затрат используется своя «валюта»: в мембране это ΔμН+ или ΔμNa+, а в цитоплазме – АТФ, креатинфосфат и другие макроэргические соединения. Непосредственным источником АТФ являются процессы субстратного и окислительного фосфорилирования. Процессы субстратного фосфорилирования наблюдаются при гликолизе и на одной из стадий цикла трикарбоновых кислот (реакция сукцинил-КоА —> сукцинат; см. главу 10). Генерация ΔμН+ и ΔμNa , используемых для окислительного фосфорилирования, осуществляется в процессе транспорта электронов в дыхательной цепи энергосопрягающих мембран.
Энергия разности потенциалов на сопрягающих мембранах может обратимо превращаться в энергию АТФ. Эти процессы катализируются Н+-АТФ-синтазой в мембранах, генерирующих протонный потенциал, или Na+-АТФ-синтазой (Na+-АТФазой) в «натриевых мембранах» алкалофильных бактерий, поддерживающих ΔμNa+ . На рис представлена схема энергетики живых клеток, использующих ΔμН+ в качестве мембранной формы конвертируемой энергии. На схеме видно, что свет или энергия субстратов дыхания утилизируется ферментами фотосинтетической или дыхательной редокс-цепи (у галобактерий – бактериородопсином). Генерируемый потенциал используется для совершения полезной работы, в частности для образования АТФ. Будучи макроэргическим соединением, АТФ выполнняет функцию аккумулирования биологической энергии и ее последующего использования для выполнения клеточных функций. «Макроэргичность» АТФ объясняется рядом особенностей его молекулы.
Химическая формула молекулы АТФ
Это прежде всего высокая плотность зарядов, сконцентрированная в «хвосте» молекулы, обеспечивающая легкость диссоциации терминального фосфата при водном гидролизе. Продукты этого гидролиза представляют собой АДФ и неорганический фосфат и далее – АМФ и неорганический фосфат. Это обеспечивает высокую величину свободной энергии гидролиза терминального фосфата АТФ в водной среде.
Взаимозаменяемость различных видов биологической энергии при выполнении клеточной работы
Красной стрелкой показана взаимозаменяемость в клетке двух клеточных видов энергии – АТФ и ΔμН+, для которых имеются также специальные буферные системы: креатинфосфат для АТФ (клетки животных) и градиент ионов Na (алкалофильные бактерии).
Тканевое дыхание и биологическое окисление. Распад органических соединений в живых тканях, сопровождающийся потреблением молекулярного кислорода и приводящий к выделению углекислого газа и воды и образованию биологических видов энергии, называется тканевым дыханием.
Тканевое дыханне – процесс поглощения тканями кислорода,
Который идет на окисление субстратов, и выделения СО2 и Н2О с образованием энергии
Тканевое дыхание представляют как конечный этап пути превращений моносахаров (в основном глюкозы) до указанных конечных продуктов, в который на разных стадиях включаются другие сахара и их производные, а также промежуточные продукты распада липидов (жирные кислоты), белков (аминокислоты) и нуклеиновых оснований. Итоговая реакция тканевого дыхания будет выглядеть следующим образом:
С6Н12О6 + 6O2 = 6СO2+ 6Н2O + 2780 кДж/моль
Схема строения дыхательной цепи во внутренней мембране митохондрий человека
http://www.youtube.com/watch?v=-_8aYKcQZ_Q&feature=related
Впервые сущность дыхания объяснил А.-Л. Лавуазье (1743-1794), обративший внимание на сходство между горением органических веществ вне организма и дыханием животных. Постепенно становились ясными принципиальные различия между этими двумя процессами: в организме окисление протекает при относительно низкой температуре в присутствии воды, и его скорость регулируется обменом веществ. В настоящее время биологическое окисление определяется как совокупность реакций окисления субстратов в живых клетках, основная функция которых – энергетическое обеспечение метаболизма. В развитие концепций биологического окисления в XX в. важнейший вклад внесли А.Н. Бах, О. Варбург, Г. Крепс, В.А. Энгельгардт, В.И. Палладин, В.А. Белицер, С.Е. Северин, В.П. Скулачев.
Потребление кислорода тканями зависит от интенсивности реакций тканевого дыхания. Наибольшей скоростью тканевого дыхания характеризуются почки, мозг, печень, наименьшей – кожа, мышечная ткань (в покое). Уравнение (2) описывает суммарный результат многоступенчатого процесса, приводящего к образованию молочной кислоты и протекающего без участия кислорода:
С6Н12Об = 2С3Н6О3 + 65 кДж/моль. (2)
Этот путь отражает, по-видимому, энергетическое обеспечение простейших форм жизни, функционировавших в бескислородных условиях. Современные анаэробные микроорганизмы (осуществляющие молочнокислое, спиртовое и уксуснокислое брожение) получают для жизнедеятельности энергию, производимую в процессе гликолиза или его модификаций.
Использование клетками кислорода открывает возможности для более полного окисления субстратов. В аэробных условиях продукты бескислородного окисления становятся субстратами цикла трикарбоновых кислот (см. главу 10), в ходе которого образуются восстановленные дыхательные переносчики НАДФН, НАДН и флавиновые коферменты. Способность НАД+ и НАДФ+ играть роль промежуточного переносчика водорода связана с наличием в их структуре амида никотиновой кислоты. При взаимодействии этих кофакторов с атомами
водорода имеет место обратимое гидрирование (присоединение атомов водорода):
Рис. Порядок расположения комплексов дыхательной цепи во внутренней мембране митохондрий
При дегидрировании субстратов образуется восстановленная форма НАД (НАДH∙H+).

Ацетоацетат → β-гидроксибутират
Дигидроксиацетон фосфат → глицерол-3-фосфат
НАДH∙H+ затем окисляется 1-м комплексом дыхательной цепи. Рассмотрим работу этого комплекса.

I комплекс цепи тканевого дыхания – НАДH∙H+-убихинон-оксидодуктаза.

Рис. Первый комплекс дыхательной цепи


II комплекс цепи тканевого дыхания – сукцинат-убихинон-оксидоредуктаза.


Окислительное фосфорилирование – один из механизмов образования АТФ

Рис. Работа «молекулярной турбины» – фактора F1. В состав F1 входит γ-субъединица, способная вращаться подобно тому, как вращается турбина в воде («молекулярная турбина»). Вращение γ-субъдиницы обуславливает конформационные изменения в αβ-протомерах. Связывание АДФ с неорганическим фосфатом происходит на участке , которая аккумулируется при генерации электро-химического потенциала и подпитывается протонным переносом через канал Fo. При вращении γ-субъединицы происходят конформационные изменения αβ-протомеров, в результате чего L-участок преобразуется в T, а T – в O. В Т-участке осуществляется синтез АТФ, а из О-участка (вследствие вращения «турбины») высвобождается АТФ. Таким образом, энергия электро-химического потенциала требуется не для синтеза АТФ, а для удаления её из активного центра V комплекса. Эта энергия расходуется также на транспорт АТФ из матрикса через митохондриальные мембраны в цитоплазму клетки, а АДФ – в обратном направлении (на работу фермента АТФ-транслоказы). Львиная доля энергии протонного потенциала расходуется на транспорт ионов кальция. Доставка субстратов тканевого дыхания также осуществляется за счёт этой энергии. Измерение «силы протонного потенциала» показало, что 1 молекула АТФ может образоваться за счёт окисления НАДН∙Н+ первым комплексом дыхательной цепи, 1 молекула АТФ – за счёт окисления убихинола третьим комплексом и 1 молекула – при окислении цитохрома с четвёртым комплексом. Именно в этих местах падение редокс-потенциала превышает 0,2 v, при таком перепаде освобождается энергия, вполне достаточная для синтеза молекулы АТФ (для синтеза АТФ необходимо затратить 42 кДж/моль). Таким образом, при окислении НАД-зависимых субстратов может образоваться 3 молекулы АТФ, а при окислении ФАД-зависимых – 2 молекулы АТФ (транспорт электронов начинается со второго комплекса дыхательной цепи). Следует заметить, что ещё в 30-х годах академик В.А. Энгельгардт обнаружил, что при тканевом дыхании на 1 молекулу кислорода расходуется 3 молекулы фосфата, т.е. отношение Р/О=3 (при использовании НАД-зависиымых субстратов тканевого дыхания). Если в среду инкубации митохондрий добавлялся сукцинат, коэффициент Р/О=2. Эти показатели были названы В.А. Белицером и Е.Т. Цыбаковой коэффициентами фосфорилирования, а I, III и IV комплексы дыхательной цепи – пунктами сопряжения (связи между процессами окисления и фосфорилирования). Согласно подсчётам известного российского биохимика В.П. Скулачёва, в сутки у человека может синтезироваться более 30 кг (!) АТФ: поскольку в сутки у человека образуется 400 мл воды (22 моля), а коэффициент Р/О=3 (66 молей АТФ), то умножая на молекулярную массу АТФ (507), получаем 707 х 66 = 33 462 (т.е.>33 кг) Транспорт электронов (окисление НАДН∙Н+ и ФАДН2 кислородом) и окислительное фосфорилирование (синтез АТФ) в норме тесно связаны Это называется сопряжением окисления и фосфорилирования. В состоянии покоя, когда окислительное фосфорилирование минимально, электрохимический градиент внутренней митохондриальной мембраны достигает величины, при которой прекращается дальнейший перенос протонов. Тем самым ингибируется транспорт электронов. Однако обнаружены вещества, например, 2,4-динитрофенол (ДНФ), которые разобщают эти процессы (ДНФ получил печальную известность как препарат для снижения массы тела). Присутствие во внутренней митохондриальной мембране агентов (ионофоров), увеличивающих их проницаемость для Н+, разобщает окислительное фосфорилирование от процесса транспорта электронов, поскольку при этом нарушается генерирование электро-химического потенциала и, следовательно, синтез АТФ. ДНФ является липофильной слабой кислотой, поэтому легко проходит через мембрану (снаружи внутрь митохондрии), ослабляя естественный ток протонов, идущий в противоположном направлении. Путь транспорта электронов, не связанный с синтезом АТФ, называется свободным, нефосфорилирующим, окислением. При свободном окислении энергия не аккумулируется, а высвобождается в виде тепла. Это имеет физиологическое значение при охлаждении организма. Частичное разобщение окисления и фосфорилирования наблюдается при многих заболеваниях, так как митохондрии являются наиболее чувствительными клеточными органеллами к действию различных повреждающих факторов. Нарушение их структуры, приводящее к частичному или полному распаду внутренней митохондриальной мембраны, неизбежно будет способствовать обратному току протонов и нарушать энергопродукцию. Поэтому особое значение приобретает стабилизация митохондриальных мембран биоантиоксидантами (витаминами Е, А и аскорбатом) при любой патологии. В ряде случаев некоторые пункты фосфорилирования могут «выключатся» – такое состояние называется разобщением окислительного фосфорилирования – и в этом случае P/O снижается: для НАД-зависимых субстратов – ниже 3; для ФАД-зависимых субстратов – ниже 2-ух. И исходя из 1-го закона термодинамики в разобщенных митохондриях увеличивается теплообразование. (Это происходит за счет того, что та энергия электронов, которая должна была быть использована для синтеза АТФ в «выключенном» пункте фосфорилирования, рассеивается в виде тепла). Процесс разобщения окислительного фосфорилирования лежит в основе лихорадки, вызванной бактериями, вирусами и другими агентами. Разобщение резко усиливается при охлаждении организма. Работа митохондрий при «выключенных» всех пунктах фосфорилирования называется сопряженной, в противном, выше описанном случае, разобщенной и дыхание при этом называется свободным. В качестве разобщителей окислительного фосфорилирования выступают слабые гидрофобные кислоты (ЖК), тиреоидные гормоны, лекарства(дикумарин, динитрофенол). ДЦ имеет механизмы шунтирования: сброс электронов и протонов по протяжению с НАД на цитохромы, или с НАД на межмембранные дегидрогеназы, на наружную мембрану и гладкую ЭПС (микросомальную цепь). Такой перенос характерен для печени. При воздействии какого-либо блокатора, возникает блок в 1-ом комплексе ДЦ, происходит накопление НАД.Н2 и становится реальной угроза гипоксии, печень может погибнуть. Чтобы этого не произошло происходит сброс НАД.Н2 с митохондрий ДЦ на микросомальную ДЦ через цитохром b5.
Регуляция процесса окислительного фосфорилирования.
Экспериментальные исследования показали, что добавление в ячейку полярографа с инкубируемыми митохондриями АДФ резко стимулировало потребление митохондриями кислорода. Накопление АДФ – регуляторный сигнал, вызывающий стимуляцию тканевого дыхания, т.е. усиление окисления субстратов тканевого дыхания. Это имеет большое физиологическое значение, так как увеличение концентрации АДФ в клетке является свидетельством низкой концентрации АТФ (соотношение адениловых нуклеотидов в клетке постоянно), следовательно, указывает на энергетическое её голодание. Отсюда вытекает и необходимость большего потребления субстратов. Помимо регуляторной роли соотношения АДФ/АТФ, в контроле скорости фосфорилирования участвуют НАД-зависимые коферменты: высокое соотношение НАДН∙Н+/НАД+, так же как и увеличение соотношения АТФ/АДФ будет снижать интенсивность тканевого дыхания. Способность генерации энергии присуща всем тканям но для того, чтобы они все зароботали, необходимо сильное охлаждение организма. Поэтому в организме есть ткань, которая обеспечивает термогенез в обычных условиях. Это бурая жировая ткань (БЖТ). Ее особенно много у новорожденных (от затылка до крестца, вдоль всей спины). У взрослого она локализуется в в определенных местах: между лопаток, в паху. Ярко выраженной способностью к теплопродукции обладает бурая жировая ткань новорождённых (а её много в организме зимнеспящих животных), которая отличается от типичной жировой ткани тем, что содержит много триацилглицеролов. Кроме того, в митохондриях этой ткани так много цитохромов, что она приобретает бурый цвет. Митохондрии бурой жировой ткани содержат разобщающий белок термогенин (димер 32 кД), который действует подобно каналу, контролирующему проводимость протонов во внутренней митохондриальной мембране. Поток протонов через этот канал ингибируется физиологическими концентрациями пуриновых нуклеотидов (АДФ, АТФ, ГДФ и др), но это ингибирование может быть устранено свободными жирными кислотами. Компоненты этой системы подчиняются гормональному контролю, в частности, норадреналину, который через систему цАМФ активирует гормончувствительную липазу, расщепляющую триацилглицеролы бурого жира с освобождением свободных жирных кислот. БЖТ имеет большое количество митохондрий, т. к. митохондрии содержат цитохромы, то это и придает этой ткани бурый цвет. Особенностью митохондрий БЖТ является отсутствие АТФ-азы, НmD+ генерируется в обычном режиме, но нет инструмента НmD(аденилаттранслаказы), трансформирующего + —> АТФ. Вместо нее НmDесть белок термогенин, который шунтирует + с наружной мембраны во НmDвнутрь и одновременно способствует рассеиванию энергии + в виде тепла, так и происходит подогрев тканей. БЖТ охватывает крупные кровеносные сосуды и согревает кровь, а потом эта кровь согревает перефирические участки тела.Это и есть несократительный термогенез. Механизм сократительного термогенеза связан с окислительным фосфорилированием. При охлаждении организма, активизируется симпатическая нервная система, в овет на это происходит выброс адреналина, под действием которого идет гликолиз, через аденилатциклазный механизм. Образующиеся при этом ЖК разобщают окислительное фосфорилирование и (согласно 1-му закону термодинамики) теплопродукция увеличивается. Именно поэтому, после принятия жирной пищи наступает состояние температурного комфорта.
Движение электронов через электронтраспортную систему дыхательных ферентов сопровождается одновременно «перекачиваем» протонов из матрикса в межмебранное пространство. Эта передислокация протонов осуществляется I, III и IV комплексами дыхательной цепи. Английский биохимик Питер Митчелл предложил рассматривать внутреннюю мембрану митохондрий как конденсатор, который со стороны матрикса заряжается отрицательно (благодаря направленному току электронов), а со стороны межмембранного пространства – положительно. Следовательно, при тканевом дыхании совершается осмотическая работа по концентрации протонов в межмембранном пространстве и возникает разность электрических потенциалов, т.е. генерируется электро-химический (или протонный) потенциал:

http://www.youtube.com/watch?v=1engJR_XWVU&feature=related
http://www.youtube.com/watch?v=_PgjsfY71AM&feature=related
Взаимное расположение компонентов дыхательной цепи с
указанием мест фосфорилирования и специфических ингибиторов
Убихинон (кофермент Q) – производное изопрена:
http://www.youtube.com/watch?v=StXlo1W3Gvg&feature=related
http://www.youtube.com/watch?v=YndC0gS3t6M&feature=related
Сопряжение цепи транспорта электронов и фосфорилирования ADP посредством протонного градиента
http://www.youtube.com/watch?v=_PgjsfY71AM&feature=related
http://www.youtube.com/watch?v=3rO26W1xG9U&feature=related
Структура компонентов комплекса I, обеспечивающего функционирование «протонной помпы» при окислении NADH
Схематическое изображение цитохромоксидазы с разрешением
0,5 нм (а) и ее активного центра с разрешением 2,8 нм (б)
НАДФН + НАД+ = НАДФ+ + НАДН + 30 кДж/моль. (4)
Перенос электронов внутри комплекса II
Ингибиторы дыхательной цепи
Изучению последовательности переноса электронов способствовало исследование действия специфических ингибиторов, блокирующих определённые этапы этого процесса (рис.).
Рис. Места действия ингибиторов ЦПЭ. Ингибиторы NADH-дегидрогеназы: ротенон – высокотоксичное вещество, содержащееся в некоторых водорослях и являющееся ядом для рыб; амитал – лекарственный препарат из группы барбитуратов. Ингибитор QH2-дегидрогеназы – антимицин А, токсичный антибиотик, продуцируемый одним из штаммов Streptomyces. Ингибиторы цитохромоксидазы – цианид, СО, H2S. Цианид наиболее токсичен для человека; он присоединяется к Fe3+ цитохромоксидазы и блокирует перенос электронов к кислороду.
Переносчики электронов, стоящие в цепи непосредственно перед блокированным этапом, становятся более восстановленными, а стоящие после этого этапа – более окисленными. Это можно обнаружить при помощи спектрофотометра, так как у окисленных и восстановленных форм переносчиков разные спектры поглощения.
РАЗОБЩЕНИЕ ДЫХАНИЯ И ФОСФОРИЛИРОВАНИЯ
Некоторые химические вещества (протонофоры) могут переносить протоны или другие ионы (ионофоры) из межмембранного пространства через мембрану в матрикс, минуя протонные каналы АТФ-синтазы.
В результате этого исчезает электрохимический потенциал и прекращается синтез АТФ. Это явление называют разобщением дыхания и фосфорилирования. В результате разобщения количество АТФ снижается, а увеличивается. В этом случае скорость окисления NADH и FADH2 возрастает, возрастает и количество поглощённого кислорода, но энергия выделяется в виде теплоты, и коэффициент Р/О резко снижается. Как правило, разобщители – липофильные вещества, легко проходящие через ли-пидныгй слой мембраны. Одно из таких веществ – 2,4-динитрофенол (рис.), легко переходящий из ионизированной формы в неионизированную, присоединяя протон в межмембранном пространстве и перенося его в матрикс.
Рис. Механизм разобщения дыхания и фос-форилирования. Протонированная форма 2,4-ди-нитрофенола переносит протоны через внутреннюю мембрану митохондрий и препятствует образованию протонного градиента.
Примерами разобщителей могут быть также некоторые лекарства, например дикумарол – антикоагулянт или метаболиты, которые образуются в организме, билирубин – продукт катаболизма гема, тироксин – гормон щитовидной железы. Все эти вещества проявляют разобщающее действие только при их высокой концентрации.
ТЕРМОРЕГУЛЯТОРНАЯ ФУНКЦИЯ ЦЕПИ ПЕРЕНОСА ЭЛЕКТРОНОВ
Hа синтез молекул АТФ расходуется примерно 40-45% всей энергии электронов, переносимых по ЦПЭ, приблизительно 25% тратится на работу по переносу веществ через мембрану. Остальная часть энергии рассеивается в виде теплоты и используется теплокровными животными на поддержание температуры тела. Кроме того, дополнительное образование теплоты может происходить при разобщении дыхания и фосфорилирования. Разобщение окислительного фосфорилирования может быть биологически полезным. Оно позволяет генерировать тепло для поддержания температуры тела у новорождённых, у зим-неспящих животных и у всех млекопитающих в процессе адаптации к холоду. У новорождённых, а также зимнеспящих животных существует особая ткань, специализирующаяся на теплопродукции посредством разобщения дыхания и фосфорилирования – бурый жир. Бурый жир содержит много митохондрий. В мембране митохондрий имеется большой избыток дыхательных ферментов по сравнению с АТФ-синтазой. Около 10% всех белков приходится на так называемый разобщающий белок (РБ-1) – термогенин. Бурый жир имеется у новорождённых, но его практически нет у взрослого человека. В последние годы появились факты, свидетельствующие о существовании в митохондриях разных органов и тканей млекопитающих разобщающих белков, похожих по своей структуре на РБ-1 бурой жировой ткани. По своей структуре термогенин близок к ATФ/AДФ-антипортеру, но не способен к транспорту нуклеотидов, хотя сохранил способность переносить анионы жирных кислот, служащих разобщителями (рис.).
Рис. Механизм разобщающего действия жирных кислот. 1- выкачивание протонов дыхательной цепью; 2 – протонирование аниона жирной кислоты; 3 – диффузия протонированной жирной кислоты к внутренней поверхности мембраны; 4 – диссоциация RCOOH с образованием RCOO– и иона Н+; 5 – перенос RCOO– посредством АТФ/АДФ-антипортера или разобщающего белка к наружной поверхности митохондри-альной мембраны.
Hа внешней стороне мембраны анион жирной кислоты присоединяет протон и в таком виде пересекает мембрану; на внутренней стороне мембраны диссоциирует, отдавая протон в матрикс и тем самым снижает протонный градиент. Образующийся анион возвращается на наружную сторону мембраны с помощью ATФ/AДФ-антипортера.
При охлаждении стимулируется освобождение норадреналина из окончаний симпатических нервов. В результате происходят активация липазы в жировой ткани и мобилизация жира из жировых депо (см. раздел 8). Образующиеся свободные жирные кислоты служат не только «топливом», но и важнейшим регулятором разобщения дыхания и фосфорилирования.
Заключительный этап катаболизма – основной источник доноров водорода для цепи переноса єлектронов
Углеводы, жирные кислоты и большинство аминокислот окисляются в конечном счёте через цикл лимонной кислоты до СО2 и Н2О. Прежде чем эти вещества вовлекаются в заключительный этап катаболизма, их углеродный скелет превращается в двухуглеродный фрагмент в форме ацетил-КоА. Именно в этой форме большая часть «топливных» молекул включается в цикл лимонной кислоты.
Ацетил-КоА образуется в специфических реакциях катаболизма жирных кислот и некоторых аминокислот. Однако главным источником ацетил-КоА служит пиро-виноградная кислота, образующаяся в реакциях катаболизма глюкозы и некоторых аминокислот.
Превращение пирувата в ацетил-КоА происходит при участии набора ферментов, структурно объединённых в пируватдегидрогеназный комплекс (ПДК). Ацетильный остаток -ацетил-КоА далее окисляется в цикле лимонной кислоты до СО2 и Н2О. В этих реакциях окисления принимают участие NAD- и FAD-зависимые дегидрогеназы, поставляющие электроны и протоны в ЦПЭ, по которой они передаются на О2.
В реакциях свободного окисления участвуют также кислород и восстановленные дыхательные переносчики (чаще всего НАДФН). Акцептором электронов является цитохром Р-450 (иногда цитохром b5). Окисление субстрата протекает по следующей схеме: SH + O2–> SOH. (5)
ГЕНЕРАЦИЯ СВОБОДНЫХ РАДИКАЛОВ В КЛЕТКЕ
Взаимопревращения свободных радикалов и их основные функции в тканях
http://www.youtube.com/watch?v=i3WZ2pyI-qM&playnext=1&list=PLD9620432F9A4D353


{kind=link}
{kind=link}
{kind=link}