БИОХИМИЧЕСКИЕ ФУНКЦИИ ПЕЧЕНИ. МЕТАБОЛИЗМ ПОРФИРИНА: ОБМЕН ЖЕЛЧНЫХ ПИГМЕНТОВ, БИОХИМИЯ ЖЕЛТУХ. БИОТРАНСФОРМАЦИЯ КСЕНОБИОТИКОВ И ЭНДОГЕННЫХ ТОКСИНОВ В ПЕЧЕНИ: МИКРОСОМАЛЬНОЕ ОКИСЛЕНИЕ, ЦИТОХРОМ Р-450
МОЧЕОБРАЗОВАТЕЛЬНАЯ ФУНКЦИЯ ПОЧЕК. ФИЗИКО-ХИМИЧЕСКИЕ СВОЙСТВА И ХИМИЧЕСКИЙ СОСТАВ НОРМАЛЬНОЙ МОЧИ. ПАТОБИОХИМИЯ ПОЧЕК. ПАТОЛОГИЧЕСКИЕ КОМПОНЕНТЫ МОЧИ
ПЕЧЕНЬ
Важнейшее значение печени в обмене веществ в первую очередь определяется тем, что она является как бы большой промежуточной станцией между портальным и общим кругом кровообращения.
http://www.youtube.com/watch?v=tat0QYxlCbo&feature=related
В печень человека более 70% крови поступает через воротную вену, остальная кровь попадает через печеночную артерию. Кровь воротной вены омывает всасывающую поверхность кишечника, и в результате большая часть веществ, всасывающихся в кишечнике, проходит через печень (кроме липидов, транспорт которых в основном осуществляется через лимфатическую систему). Таким образом, печень функционирует как первичный регулятор содержания в крови веществ, поступающих в организм с пищей. Доказательством справедливости данного положения является следующий общий факт: несмотря на то что всасывание питательных веществ из кишечника в кровь происходит прерывисто, непостоянно, в связи с чем в портальном круге кровообращения могут наблюдаться изменения концентрации ряда веществ (глюкоза, аминокислоты и др.), в общем круге кровообращения изменения в концентрации указанных соединений незначительны. Все это подтверждает важную роль печени в поддержании постоянства внутренней среды организма. Печень выполняет также крайне важную экскреторную функцию, теснейшим образом связанную с ее детоксикационной функцией. В целом без преувеличения можно констатировать, что в организме нет путей обмена веществ, которые прямо или косвенно не контролировались бы печенью, в связи с чем многие важнейшие функции печени уже рассматривались в соответствующих главах учебника. В данной главе будет сделана попытка дать обобщающие представления о роли печени в обмене веществ целостного организма.
ХИМИЧЕСКИЙ СОСТАВ ПЕЧЕНИ
У взрослого здорового человека масса печени составляет в среднем
В печени может содержаться 150–200 г гликогена. Как правило, при тяжелых паренхиматозных поражениях печени количество гликогена в ней уменьшается. Напротив, при некоторых гликогенозах содержание гликогена достигает 20% и более от массы печени.
Разнообразен и минеральный состав печени. Количество железа, меди, марганца, никеля и некоторых других элементов превышает их содержание в других органах и тканях.
ПЕЧЕНИ В УГЛЕВОДНОМ ОБМЕНЕ
Основная роль печени в углеводном обмене заключается в обеспечении постоянства концентрации глюкозы в крови. Это достигается регуляцией между синтезом и распадом гликогена, депонируемого в печени.
В печени синтез гликогена и его регуляция в основном аналогичны тем процессам, которые протекают в других органах и тканях, в частности в мышечной ткани. Синтез гликогена из глюкозы обеспечивает в норме временный резерв углеводов, необходимый для поддержания концентрации глюкозы в крови в тех случаях, если ее содержание значительно уменьшается (например, у человека это происходит при недостаточном поступлении углеводов с пищей или в период ночного «голодания»).

Синтез и распад гликогена
http://www.youtube.com/watch?v=p-lFJVOkFwk
Необходимо подчеркнуть важную роль фермента глюкокиназы в процессе утилизации глюкозы печенью. Глюкокиназа, подобно гексокиназе, катализирует фосфорилирование глюкозы с образованием глюкозо-6-фосфата, при этом активность глюкокиназы в печени почти в 10 раз превышает активность гексокиназы.
http://www.youtube.com/watch?v=O5eMW4b29rg&feature=related
Важное различие между этими двумя ферментами заключается в том, что глюкокиназа в противоположность гексокиназе имеет высокое значение КМ для глюкозы и не ингибируется глюкозо-6-фосфатом.
Синтез гликогена
Распад гликогена
После приема пищи содержание глюкозы в воротной вене резко возрастает: в тех же пределах увеличивается и ее внутрипеченочная концентрация . Повышение концентрации глюкозы в печени вызывает существенное увеличение активности глюкокиназы и автоматически увеличивает поглощение глюкозы печенью (образовавшийся глюкозо-6-фосфат либо затрачивается на синтез гликогена, либо расщепляется).
|
Механизм действия фосфорилазы гликогена |
Особенности метаболизма гликогена в печени и мышцах
http://www.youtube.com/watch?v=oBL0OC3IavI
Считают, что основная роль печени – расщепление глюкозы – сводится прежде всего к запасанию метаболитов-предшественников, необходимых для биосинтеза жирных кислот и глицерина, и в меньшей степени к окислению ее до СО2 и Н2О. Синтезированные в печени триглицериды в норме выделяются в кровь в составе липопротеинов и транспортируются в жировую ткань для более «постоянного» хранения.
В реакциях пентозофосфатного пути в печени образуется НАДФН, используемый для восстановительных реакций в процессах синтеза жирных кислот, холестерина и других стероидов. Кроме того, при этом образуются пентозофосфаты, необходимые для синтеза нуклеиновых кислот.
Пентозофосфатный путь превращения глюкозы
Наряду с утилизацией глюкозы в печени происходит и ее образование. Непосредственным источником глюкозы в печени служит гликоген. Распад гликогена в печени происходит в основном фосфоролитическим путем. В регуляции скорости гликогенолиза в печени большое значение имеет система циклических нуклеотидов. Кроме того, глюкоза в печени образуется также в процессе глюконеогенеза.
Глюконеогенез. Ферменты: 1- пируваткарбоксилаза, 2- фосфоенолпируваткарбоксикиназа, 3- фосфатаза фру-1,6-дифосфата, 4- глюкозо-6-фосфатаза.
Основными субстратами глюконеогенеза служат лактат, глицерин и аминокислоты. Принято считать, что почти все аминокислоты, за исключением лейцина, могут пополнять пул предшественников глюконеогенеза.
При оценке углеводной функции печени необходимо иметь в виду, что соотношение между процессами утилизации и образования глюкозы регулируется прежде всего нейрогуморальным путем при участии желез внутренней секреции.
Центральную роль в превращениях глюкозы и саморегуляции углеводного обмена в печени играет глюкозо-6-фосфат. Он резко тормозит фосфоролитическое расщепление гликогена, активирует ферментативный перенос глюкозы с уридиндифосфоглюкозы на молекулу синтезирующегося гликогена, является субстратом для дальнейших гликолитических превращений, а также окисления глюкозы, в том числе по пентозофосфатному пути. Наконец, расщепление глюкозо-6-фосфата фосфатазой обеспечивает поступление в кровь свободной глюкозы, доставляемой током крови во все органы и ткани (рис. ).
Рис. Участие глюкозо-6-фосфата в метаболизме углеводов.
Рис. Гормональная регуляция системы фруктозо-2,6-бисфосфата (Ф-2,6-Р2) в печени при участии цАМФ-зависимых протеинкиназ.
http://www.youtube.com/watch?v=oBL0OC3IavI
Как отмечалось, наиболее мощным аллостерическим активатором фосфофруктокиназы-1 и ингибитором фруктозо-1,6-бисфосфатазы печени является фруктозо-2,6-бисфосфат (Ф-2,6-Р2). Повышение в гепатоцитах уровня Ф-2,6-Р2 способствует усилению гликолиза и уменьшению скорости глюконеогенеза. Ф-2,6-Р2 снижает ингибирующее действие АТФ на фосфо-фруктокиназу-1 и увеличивает сродство этого фермента к фруктозо-6-фосфату. При ингибировании фруктозо-1,6-бисфосфатазы Ф-2,6-Р2 возрастает значение КМ для фруктозо-1,6-бисфосфата. Содержание Ф-2,6-Р2 в печени, сердце, скелетной мускулатуре и других тканях контролируется бифункциональным ферментом, который осуществляет синтез Ф-2,6-Р2 из фруктозо-6-фосфата и АТФ и гидролиз его до фруктозо-6-фосфата и Pi, т.е. фермент одновременно обладает и киназной, и бисфосфатазной активностью. Бифункциональный фермент (фосфофруктокиназа-2/фруктозо-2,6-бисфосфатаза), выделенный из печени крысы, состоит из двух идентичных субъединиц с мол. массой 55000, каждая из которых имеет два различных каталитических центра. Киназный домен при этом расположен на N-конце, а бисфосфатазный – на С-конце каждой из полипептидных цепей. Известно также, что бифункциональный фермент печени является прекрасным субстратом для цАМФ-зависимой протеинкиназы А. Под действием протеинкиназы А происходит фосфорилирование остатков серина в каждой из субъединиц бифункционального фермента, что приводит к снижению его киназной и повышению бисфосфатазной активности. Заметим, что в регуляции активности бифункционального фермента существенная роль принадлежит гормонам, в частности глюкагону.
При многих патологических состояниях, в частности при сахарном диабете, отмечаются существенные изменения в функционировании и регуляции системы Ф-2,6-Р2. Установлено, что при экспериментальном (стептозотоциновом) диабете у крыс на фоне резкого увеличения уровня глюкозы в крови и моче в гепатоцитах содержание Ф-2,6-Р2 снижено. Следовательно, снижается скорость гликолиза и усиливается глюконео-генез. Данный факт имеет свое объяснение. Возникающие у крыс при диабете нарушения гормонального фона: увеличение концентрации глю-кагона и уменьшение содержания инсулина – обусловливают повышение концентрации цАМФ в ткани печени, усиление цАМФ-зависимого фосфорилирования бифункционального фермента, что в свою очередь приводит к снижению его киназной и повышению бисфосфатазной активности. Таков может быть механизм снижения уровня Ф-2,6-Р2 в гепатоцитах при экспериментальном диабете. По-видимому, существуют и другие механизмы, ведущие к снижению уровня Ф-2,6-Р2 в гепатоцитах при стрептозото-циновом диабете. Показано, что при экспериментальном диабете в ткани печени имеет место снижение активности глюкокиназы (возможно, и снижение количества данного фермента). Это приводит к падению скорости фосфорилирования глюкозы, а затем к снижению содержания фруктозо-6-фосфата – субстрата бифункционального фермента. Наконец, в последние годы было показано, что при стрептозотоциновом диабете уменьшается количество мРНК бифункционального фермента в гепатоцитах и как следствие – снижается уровень Ф-2,6-Р2 в ткани печени, усиливается глюко-неогенез. Все это еще раз подтверждает положение, что Ф-2,6-Р2, являясь важным компонентом в цепи передачи гормонального сигнала, выступает в роли третичного посредника при действии гормонов, прежде всего на процессы гликолиза и глюконеогенеза.
Рассматривая промежуточный обмен углеводов в печени, необходимо также остановиться на превращениях фруктозы и галактозы. Поступающая в печень фруктоза может фосфорилироваться в положении 6 до фруктозо-6-фосфата под действием гексокиназы, обладающей относительной специфичностью и катализирующей фосфорилирование, кроме глюкозы и фруктозы, еще и маннозы. Однако в печени существует и другой путь: фруктоза способна фосфорилироваться при участии более специфического фермента – фруктокиназы. В результате образуется фруктозо-1-фосфат. Эта реакция не блокируется глюкозой. Далее фруктозо-1-фосфат под действием альдолазы расщепляется на две триозы: диоксиацетонфосфат и глицераль-дегид. Под влиянием соответствующей киназы (триокиназы) и при участии АТФ глицеральдегид подвергается фосфорилированию до глицеральдегид-3-фосфата. Последний (в него легко переходит и диоксиацетонфосфат) подвергается обычным превращениям, в том числе с образованием в качестве промежуточного продукта пировиноградной кислоты.
Связь пентозофосфатного пути превращения глюкозы с гликолизом и глюконеогенезом
Следует отметить, что при генетически обусловленной нетолерантности к фруктозе или недостаточной активности фруктозо-1,6-бисфосфатазы наблюдается индуцируемая фруктозой гипогликемия, возникающая вопреки наличию больших запасов гликогена. Вероятно, фруктозо-1-фосфат и фруктозо-1,6-бисфосфат ингибируют фосфорилазу печени по аллосте-рическому механизму.
Известно также, что метаболизм фруктозы по гликолитическому пути в печени происходит гораздо быстрее, чем метаболизм глюкозы. Для метаболизма глюкозы характерна стадия, катализируемая фосфофрукто-киназой-1. Как известно, на этой стадии осуществляется метаболический контроль скорости катаболизма глюкозы. Фруктоза минует эту стадию, что позволяет ей интенсифицировать в печени процессы метаболизма, ведущие к синтезу жирных кислот, их эстерификацию и секрецию липопротеинов очень низкой плотности; в результате может увеличиваться концентрация триглицеридов в плазме крови.
Галактоза в печени сначала фосфорилируется при участии АТФ и фермента галактокиназы с образованием галактозо-1-фосфата. Для га-лактокиназы печени плода и ребенка характерны значения КМ и Vмaкс, примерно в 5 раз превосходящие таковые у ферментов взрослого человека. Большая часть галактозо-1-фосфата в печени превращается в ходе реакции, катализируемой гексозо-1-фосфат-уридилилтрансферазой:
УДФ-глюкоза + Галактозо-1-фосфат –> УДФ-галактоза + Глюкозо-1-фосфат.
Это уникальная трансферазная реакция возвращения галактозы в основное русло углеводного метаболизма. Наследственная утрата гексозо-1-фосфат-уридилилтрансферазы приводит к галактоземии – заболеванию, для которого характерны умственная отсталость и катаракта хрусталика. В этом случае печень новорожденных теряет способность метаболизи-ровать D-галактозу, входящую в состав лактозы молока.
РЕГУЛЯЦИЯ ОБМЕНА ОСНОВНЫХ ЭНЕРГОНОСИТЕЛЕЙ ПРИ НОРМАЛЬНОМ РИТМЕ ПИТАНИЯ
1. Энергетическая ценность основных пищевых веществ выражается в килокалориях и составляет: для углеводов – 4 ккал/г, для жиров – 9 ккал/г, для белков – 4 ккал/г. Взрослому здоровому человеку в сутки требуется 2000- 3000 ккал (8000-12 000 кДж) энергии.
При обычном ритме питания промежутки между приемами пищи составляют 4-5 часов с 8-12-часовым ночным перерывом. В течение пищеварения и абсорбтивного периода (2-4 часа) основные энергоносители, используемые тканями (глюкоза, жирные кислоты, аминокислоты), могут поступать в кровь непосредственно из пищеварительного тракта. В постабсорбтивном периоде (промежуток времени после завершения пищеварения до следующего приема пищи) и при голодании энергетические субстраты образуются в процессе катаболизма депонированных энергоносителей. Основную роль в регуляции этих процессов играют инсулин и глюкагон. Антагонистами инсулина являются также адреналин, кортизол, иодтиронины и соматотропин (так называемые контринсулярные гормоны).
Инсулин и контринсулярные гормоны обеспечивают баланс между потребностями и возможностями организма в получении энергии, необходимой для нормального функционирования и роста. Этот баланс определяется как энергетический гомеостаз. При нормальном ритме питания концентрация глюкозы в крови поддерживается на уровне 65-110 мг/дл (3,58-6,05 ммоль/л) благодаря влиянию двух основных гормонов – инсулина и глюкагона. Инсулин и глюкагон – главные регуляторы метаболизма при смене состояний пищеварения, постабсорбтивного периода и голодания. На периоды пищеварения приходится 10-15 час в сутки, а расход энергии происходит в течение 24 часов. Поэтому часть энергоносителей во время пищеварения запасается для использования в постабсорбтивном периоде.
Печень, жировая ткань и мышцы – главные органы, обеспечивающие изменения метаболизма в соответствии с ритмом питания. Режим запасания включается после приема пищи и сменяется режимом мобилизации запасов после завершения абсорбтивного периода.
2. Изменения метаболизма основных энергоносителей в абсорбтивном периоде обусловлены, в основном, высоким инсулин-глюкагоновым индексом (рис.).
Рис. Пути использования основных энергоносителей в абсорбтивном периоде: 1 – биосинтез гликогена в печени; 2 – гликолиз; 3 – биосинтез ТАГ в печени; 4 – биосинтез ТАГ в жировой ткани; 5 – биосинтез гликогена в мышцах; 6 – биосинтез белков в разных тканях, в том числе в печени; ЖК – жирные кислоты
Изменения метаболизма в печени. В печени увеличивается потребление глюкозы, что является следствием ускорения метаболических путей, в которых глюкоза превращается в депонируемые формы энергоносителей: гликоген и жиры.
При повышении концентрации глюкозы в гепатоцитах происходит активация глюкокиназы, превращающей глюкозу в глюкозо-6-фосфат. Кроме этого, инсулин индуцирует синтез мРНК глюкокиназы. В результате повышается концентрация глюкозо-6-фосфата в гепатоцитах, что обусловливает ускорение синтеза гликогена. Этому также способствует одновременная инактивация гликогенфосфорилазы и активация гликогенсинтазы. Под влиянием инсулина в гепатоцитах ускоряется гликолиз в результате повышения активности и количества ключевых ферментов: глюкокиназы, фосфофруктокиназы и пируваткиназы. В то же время происходит торможение глюконеогенеза в результате инактивации фруктозо-1,6-бисфосфатазы и репрессии инсулином синтеза фосфоенолпируваткарбоксикиназы – ключевых ферментов глюконеогенеза.
Повышение концентрации глюкозо-6-фосфата в гепатоцитах в абсорбтивном периоде сочетается с активным использованием NADPH для синтеза жирных кислот, что способствует стимуляции пентозофосфатного пути.
Ускорение синтеза жирных кислот обеспечивается доступностью субстратов (ацетил-КоА и NADPH), образующихся при метаболизме глюкозы, а также активацией и индукцией ключевых ферментов синтеза жирных кислот инсулином.
Аминокислоты, поступающие в печень из пищеварительного тракта, используются для синтеза белков и других азотсодержащих соединений, а их излишек либо поступает в кровь и транспортируется в другие ткани, либо дезаминируется с последующим включением безазотистых остатков в общий путь катаболизма.
Изменения метаболизма в адипоцитах. Основная функция жировой ткани – запасание энергоносителей в форме триацилглицеролов. Под влиянием инсулина ускоряется транспорт глюкозы в адипоциты. Повышение внутриклеточной концентрации глюкозы и активация ключевых ферментов гликолиза обеспечивают образование ацетил-КоА и глицерол-3-фосфата, необходимых для синтеза ТАГ. Стимуляция пентозофосфатного пути обеспечивает образование NADPH, необходимого для синтеза жирных кислот. Однако биосинтез жирных кислот de novo в жировой ткани человека протекает с высокой скоростью только после предшествующего голодания. При нормальном ритме питания для синтеза ТАГ используются в основном жирные кислоты, поступающие из хиломикронов и ЛПОНП под действием ЛП-липазы.
Так как гормончувствительная ТАГ-липаза в абсорбтивном состоянии находится в дефосфорилированной, неактивной форме, процесс липолиза тормозится.
Изменение метаболизма в мышцах. Под влиянием инсулина ускоряется транспорт глюкозы в клетки мышечной ткани. Глюкоза фосфорилируется и окисляется для обеспечения клеток энергией, а также используется для синтеза гликогена. Жирные кислоты, поступающие из хиломикронов и ЛПОНП, в этот период играют незначительную роль в энергетическом обмене мышц. Поток аминокислот в мышцы и биосинтез белков также возрастает под влиянием инсулина, особенно после приема белковой пищи и в период мышечной работы.
3. Изменения метаболизма основных энергоносителей при смене абсорбтивного состояния на постабсорбтивное. В постабсорбтивном периоде при снижении инсулин-глюкагонового индекса изменения метаболизма направлены главным образом на поддержание концентрации в крови глюкозы, которая служит главным энергетическим субстратом для мозга и единственным источником энергии для эритроцитов. Основные изменения метаболизма в этот период происходят в печени и жировой ткани и направлены на пополнение глюкозы за счет внутренних резервов и на использование других энергетических субстратов (жиров и аминокислот) (рис.).
Рис. Пути использования основных энергоносителей при смене абсорбтивного состояния на постабсорбтивное: I – снижение инсулин-глюкагонового индекса; 2 – распад гликогена; 3, 4 – транспорт глюкозы в мозг и эритроциты; 5 – катаболизм жиров; 6 – транспорт жиров в печень и мышцы; 7 – синтез кетоновых тел в печени; 8 – транспорт кетоновых тел в мышцы; 9 – глюконеогенез из аминокислот; 10 – синтез и выведение мочевины; II – транспорт лактата в печень и включение в глюконеогенез; 12 – глюконеогенез из глицерола; КТ -кетоновые тела; ЖК – жирные кислоты
Изменения метаболизма в печени. Под влиянием глюкагона ускоряется мобилизация гликогена. Запасы гликогена в печени истощаются в течение 18-24-часового голодания. Главным источником глюкозы по мере исчерпания запасов гликогена становится глюконеогенез, который начинает ускоряться через 4-6 часов после последнего приема пищи. Субстратами для синтеза глюкозы служат лактат, глицерол и аминокислоты. Скорость синтеза жирных кислот снижается вследствие фосфорилирования и инактивации ацетил-КоА-карбоксилазы при фосфорилировании, а скорость β-окисления возрастает. Вместе с тем увеличивается снабжение печени жирными кислотами, которые транспортируются из жировых депо в результате ускорения липолиза. Ацетил-КоА, образующийся при окислении жирных кислот, используется в печени для синтеза кетоновых тел.
В жировой ткани снижается скорость синтеза ТАГ и стимулируется липолиз. Стимуляция липолиза является результатом активации гормончувствительной ТАГ-липазы адипоцитов под влиянием глюкагона. Жирные кислоты становятся важными источниками энергиии в печени, мышцах и жировой ткани.
Таким образом, в постабсорбтивном периоде концентрация глюкозы в крови поддерживается на уровне 60-100 мг/дл (3,5-5,5 ммоль/л), а уровень жирных кислот и кетоновых тел возрастает.
РОЛЬ ПЕЧЕНИ В ЛИПИДНОМ ОБМЕНЕ
Рис. . Участие печени в жировом обмене
Рис. Основные этапы метаболизма липидов в гепатоците
http://www.youtube.com/watch?v=5HF9itMaOKo
Ферментные системы печени способны катализировать все реакции или значительное большинство реакций метаболизма липидов. Совокупность этих реакций лежит в основе таких процессов, как синтез высших жирных кислот, триглицеридов, фосфолипидов, холестерина и его эфиров, а также липолиз триглицеридов, окисление жирных кислот, образование ацетоновых (кетоновых) тел и т.д. Напомним, что ферментативные реакции синтеза триглицеридов в печени и жировой ткани сходны. Так, КоА-производные жирной кислоты с длинной цепью взаимодействуют с глицерол-3-фосфатом с образованием фосфатидной кислоты, которая затем гидролизуется до диглицерида. Путем присоединения к последнему еще одной молекулы КоА-производного жирной кислоты образуется триглицерид. Синтезированные в печени три-глицериды либо остаются в печени, либо секретируются в кровь в форме липопротеинов. Секреция происходит с известной задержкой (у человека 1–3 ч). Задержка секреции, вероятно, соответствует времени, необходимому для образования липопротеинов. Основным местом образования плазменных пре-β-липопротеинов (липопротеины очень низкой плотности – ЛПОНП) и α-липопротеинов (липопротеины высокой плотности – ЛПВП) является печень.
ХС – холестерин
ТГ – триглицериды
ЖК – жирные кислоты
ХМ – хиломикроны
ЛПЛ – липопротеинлипаза
ТГЛ – триглицеридлипаза.
http://www.youtube.com/watch?v=97uiV4RiSAY
Рассмотрим образование ЛПОНП. Согласно данным литературы, основной белок апопротеин В-100 (апо Б-100) липопротеинов синтезируется в рибосомах шероховатого эндоплазматического ретикулума гепатоцитов. В гладком эндоплазматическом ретикулуме, где синтезируются и липидные компоненты, происходит сборка ЛПОНП. Одним из основных стимулов образования ЛПОНП является повышение концентрации неэстерифици-рованных жирных кислот (НЭЖК). Последние либо поступают в печень с током крови, будучи связанными с альбумином, либо синтезируются непосредственно в печени. НЭЖК служат главным источником образования триглицеридов (ТГ). Информация о наличии НЭЖК и ТГ передается на мембранно-связанные рибосомы шероховатого эндоплазматического ретикулума, что в свою очередь является сигналом для синтеза белка (апо В-100). Синтезированный белок внедряется в мембрану шероховатого ретикулума, и после взаимодействия с фосфолипидным бислоем от мембраны отделяется участок, состоящий из фосфолипидов (ФЛ) и белка, который и является предшественником ЛП-частицы. Далее белокфосфо-липидный комплекс поступает в гладкий эндоплазматический ретикулум, где взаимодействует с ТГ и эстерифицированным холестерином (ЭХС), в результате чего после соответствующих структурных перестроек формируются насцентные, т.е. незавершенные, частицы (н-ЛПОНП). Последние поступают через тубулярную сеть аппарата Гольджи в секреторные везикулы и в их составе доставляются к поверхности клетки, после чего очень низкой плотности (ЛПОНП) в печеночной клетке (по А.Н. Климову и Н.Г. Никульчевой) путем экзоцитоза выделяются в перисинусоидные пространства (пространства Диссе). Из последнего н-ЛПОНП поступают в просвет кровяного синусоида, где происходят перенос апопротеинов С из ЛПВП на н-ЛПОНП и достраивание последних (рис. 16.3). Установлено, что время синтеза апо В-100, образования липид-белковых комплексов и секреции готовых частиц ЛПОНП составляет 40 мин.
Рис. Образование липопротеинов
1 – ядро; 2 – шероховатый эндоплазматический ретикулум; 3 – гладкий эндоплазматический ре-тикулум, синтезированные в нем липиды и образовавшиеся н-ЛПОНП; 4 – аппарат Гольджи; 5 -секреторная везикула с частицей н-ЛПОНП; 6 -частица с н-ЛПОНП в пространстве Диссе; 7 -перенос апопротеинов С с ЛПВП на н-ЛПОНП; 8 – частица нативных ЛПОНП.
http://www.youtube.com/watch?v=x-4ZQaiZry8
У человека основная масса β-липопротеинов (липопротеины низкой плотности – ЛПНП) образуется в плазме крови из ЛПОНП при действии липопротеинлипазы. В ходе этого процесса образуются сначала промежуточные короткоживущие липопротеины (Пр.ЛП), а затем формируются частицы, обедненные триглицеридами и обогащенные холестерином, т.е. ЛПНП.
При высоком содержании жирных кислот в плазме их поглощение печенью возрастает, усиливается синтез триглицеридов, а также окисление жирных кислот, что может привести к повышенному образованию кетоновых тел.
Следует подчеркнуть, что кетоновые тела образуются в печени в ходе так называемого β-гидрокси-β-метилглутарил-КоА пути. Однако существует мнение, что ацетоацетил-КоА, являющийся исходным соединением при кетогенезе, может образоваться как непосредственно в ходе β-окисле-ния жирных кислот, так и в результате конденсации ацетил-КоА [Марри Р. и др., 1993]. Из печени кетоновые тела током крови доставляются в ткани и органы (мышцы, почки, мозг и др.), где они быстро окисляются при участии соответствующих ферментов, т.е. по сравнению с другими тканями печень является исключением.
В печени происходит интенсивный распад фосфолипидов, а также их синтез. Помимо глицерина и жирных кислот, которые входят в состав нейтральных жиров, для синтеза фосфолипидов необходимы неорганические фосфаты и азотистые соединения, в частности холин, для синтеза фосфатидхолина. Неорганические фосфаты в печени имеются в достаточном количестве. При недостаточном образовании или недостаточном поступлении в печень холина синтез фосфолипидов из компонентов нейтрального жира становится либо невозможным, либо резко снижается и нейтральный жир откладывается в печени. В этом случае говорят о жировой инфильтрации печени, которая может затем перейти в ее жировую дистрофию. Иными словами, синтез фосфолипидов лимитируется количеством азотистых оснований, т.е. для синтеза фосфоглицеридов необходим либо холин, либо соединения, которые могут являться донорами метильных групп и участвовать в образовании холина (например, метионин). Такие соединения получили название липотропных веществ. Отсюда становится ясным, почему при жировой инфильтрации печени весьма полезен творог, содержащий белок казеин, в составе которого имеется большое количество остатков аминокислоты метионина.
Рассмотрим роль печени в обмене стероидов, в частности холестерина. Часть холестерина поступает в организм с пищей, но значительно большее количество его синтезируется в печени из ацетил-КоА. Биосинтез холестерина в печени подавляется экзогенным холестерином, т.е. получаемым с пищей.
Таким образом, биосинтез холестерина в печени регулируется по принципу отрицательной обратной связи. Чем больше холестерина поступает с пищей, тем меньше его синтезируется в печени, и наоборот. Принято считать, что действие экзогенного холестерина на биосинтез его в печени связано с торможением β-гидрокси-β-метилглутарил-КоА-редуктазной реакции:
Часть синтезированного в печени холестерина выделяется из организма вместе с желчью, другая часть превращается в желчные кислоты и используется в других органах для синтеза стероидных гормонов и иных соединений.
В печени холестерин может взаимодействовать с жирными кислотами (в виде ацил-КоА) с образованием эфиров холестерина. Синтезированные в печени эфиры холестерина поступают в кровь, в которой содержится также определенное количество свободного холестерина.
РОЛЬ ПЕЧЕНИ В ОБМЕНЕ БЕЛКОВ
Печень играет центральную роль в обмене белков. Она выполняет следующие основные функции: синтез специфических белков плазмы; образование мочевины и мочевой кислоты; синтез холина и креатина; трансаминирование и дезаминирование аминокислот, что весьма важно для взаимных превращений аминокислот, а также для процесса глюконеогенеза и образования кетоновых тел. Все альбумины плазмы, 75–90% α-глобу-линов и 50% β-глобулинов синтезируются гепатоцитами. Лишь γ-гло-булины продуцируются не гепатоцитами, а системой макрофагов, к которой относятся звездчатые ретикулоэндотелиоциты (клетки Купфера). В основном γ-глобулины образуются в печени. Печень является единственным органом, где синтезируются такие важные для организма белки, как протромбин, фибриноген, проконвертин и проакцелерин.
При заболеваниях печени определение фракционного состава белков плазмы (или сыворотки) крови нередко представляет интерес как в диагностическом, так и в прогностическом плане. Известно, что патологический процесс в гепатоцитах резко снижает их синтетические возможности. В результате содержание альбумина в плазме крови резко падает, что может привести к снижению онкотического давления плазмы крови, развитию отеков, а затем асцита. Отмечено, что при циррозах печени, протекающих с явлениями асцита, содержание альбуминов в сыворотке крови на 20% ниже, чем при циррозах без асцита.
http://www.youtube.com/watch?v=nXRWkorYFXc
Нарушение синтеза ряда белковых факторов системы свертывания крови
при тяжелых заболеваниях печени может привести к геморрагическим явлениям.
Детоксикация различных веществ в печени
http://www.youtube.com/watch?v=3DgxjDalZW0
http://www.youtube.com/watch?v=7ON7oMvNehg&feature=related
Роль печени в пигментном обмене
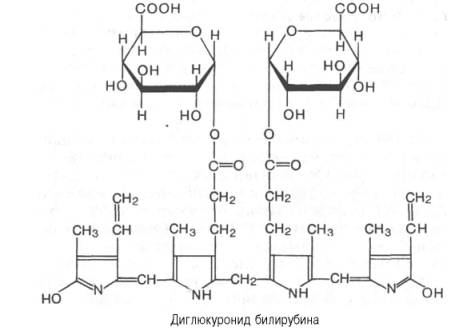
Рис. Нормальный обмен уробилиногеновых тел (схема).
ЖЕЛЧЬ
Распад гемоглобина в тканях (образование желчных пигментов)
http://www.youtube.com/watch?v=nXRWkorYFXc
|
|


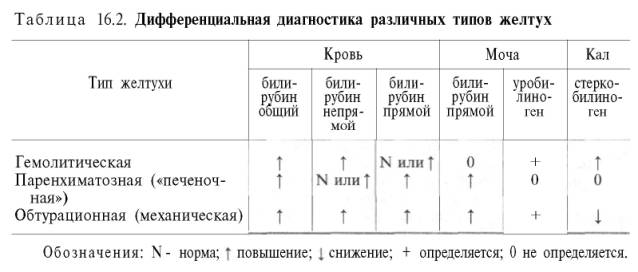
Основные лабораторные признаки желтух различного происхождения
|
|

МЕХАНИЗМЫ ОБЕЗВРЕЖИВАНИЯ КСЕНОБИОТИКОВ
Таблица 12-1. Возможные модификации ксенобиотиков в первой фазе обезвреживания
|
|
|
|
|

В метаболизме ксенобиотиков могут принимать участие ферменты почек, лёгких, кожи и ЖКТ, но наиболее активны они в печени. К группе микросомальных ферментов относят специфические оксидазы, различные гидролазы и ферменты конъюгации.
1. Основные ферменты микросомальных электронтранспортных цепей



Рис. 12-2. Электронтранспортные цепи ЭР. RH – субстрат цитохрома Р450; стрелками показаны реакции переноса электронов. В одной системе NADPH окисляется NADPH цитохром Р450-редуктазой, которая затем передаёт электроны на целое семейство цитохромов Р450. Вторая сисгема включает в себя окисление NADH цитохром b5-редуктазой, электроны переходят на цитохром b5; восстановленную форму цитохрома b5 окисляет стеароил-КоА-десатураза, которая переносит электроны на О2. Один атом кислорода принимает 2 е и переходит в форму О2-. Донором электронов служит NADPH, который окисляется NADPH-цитохром Р450 редуктазой. О2- взаимодействует с протонами: О2- + 2Н+ → Н2О, и образуется вода. Второй атом молекулы кислорода включается в субстрат RH, образуя гидроксильную группу вещества R-OH.
2. Функционирование цитохрома Р450
Суммарное уравнение реакции гидроксилирования вещества RH ферментами микросомального окисления:
RH + О2 + NADPH + Н+ → ROH + Н2О + NADP+ .
3. Свойства системы микросомальногоокисления
Широкая субстратная специфичность. Изоформы Р450

Рис. 12-3. Транспорт электронов при монооксигеназном окислении с участием Р450. Связывание (1) в активном центре цитохрома Р450 вещества RH активирует восстановление железа в теме – присоединяется первый электрон (2). Изменение валентности железа увеличивает сродство комплекса P450-Fe2+·RH к молекуле кислорода (3). Появление в центре связывания цитохрома Р450 молекулы О2 ускоряет присоединение второго электрона и образование комплекса P450-Fe2+O2–-RH (4). На следующем этапе (5) Fe2+ окисляется, второй электрон присоединяется к молекуле кислорода P450-Fe3+O22-. Восстановленный атом кислорода (О2-) связывает 2 протона, и образуется 1 молекула воды. Второй атом кислорода идёт на построение ОН-группы (6). Модифицированное вещество R-OH отделяется от фермента (7).
Регуляция активности микросомальной системы окисления
Б. Конъюгация – вторая фаза обезвреживание веществ
1. Участие трансферам в реакциях конъюгации

Рис. Уридиндифосфоглюкуроновая кислота (УДФ-C6H9O6).
В общем виде реакция с участием УДФ-глюкуронилтрансферазы записывается так:
ROH + УДФ-С6Н9О6 = RO-C6H9O6 + УДФ.

Рис. 3′-Фосфоаденозин-5′-фосфосульфат(ФАФ-SО3Н).
Таблица 12-2. Основные ферменты и метаболиты, участвующие в конъюгации
Реакция с участием сульфотрансферазы в общем виде записывается так:
ROH + ФАФ-SO3H = RO-SO3H + ФАФ.
· путём конъюгации субстрата R с глутатионом (GSH):
· в результате нуклеофильного замещения:
· восстановления органических пероксидов до спиртов:
R-HC-O-OH + 2 GSH → R-HC-OH + GSSG + H2O
· В реакции: ООН – гидропероксидная группа, GSSG – окисленный глутатион.


Рис. Oбезвреживание 1-хлор, 2,4-динитробензола с участием глутатиона.
Ацетилтрансферазы, метилтрансферазы
2. Роль эпоксидгидролаз в образовании диолов

Рис. Обезвреживание бензантрацена. Е1 – фермент микросомальной системы; Е2 – эпоксидгидратаза.
Гниение аминокислот в кишечнике. Обезвреживание и выведение продуктов гниения из организма
Образование и обезвреживание n-крезола и фенола

Рис. Катаболизм тирозина под действием бактерий. E – бактериальные ферменты.

Рис. Конъюгация фенола и крезола с ФАФС. E – сульфотрансфераза.

Образование и обезвреживание индола и скатола

Рис. Катаболизм триптофана под действием бактерий. E – бактериальные ферменты.

Рис. Участие сульфотрансферазы в обезвреживании индола. E – сульфотрансфераза.
Обезвреживание бензойной кислоты

Рис. Образование гиппуровой кислоты из бензойной кислоты и глицина. E-глицинтрансфераза
Скорость этой реакции отражает функциональное состояние печени.
Связывание, транспорт и выведение ксенобиотиков
Участие Р-гликопротеина в выведении ксенобиотиков

В норме его функция состоит в экскреции ионов хлора и гидрофобных токсичных соединений из клеток.

При химиотерапии злокачественных процессов начальная эффективность лекарства часто постепенно падает. Более того, развивается множественная лекарственная устойчивость, т.е. устойчивость не только к этому лечебному препарату, но и целому ряду других лекарств. Это происходит потому, что противоопухолевые лекарства индуцируют синтез Р-гликопротеина, глутатионтрансферазы и глутатиона. Использование веществ, ингибирующих или активирующих синтез Р-гликопротеина, а также ферменты синтеза глутатиона, повышает эффективность химиотерапии.
МОЧЕОБРАЗОВАТЕЛЬНАЯ ФУНКЦИЯ ПОЧЕК. ФИЗИКО-ХИМИЧЕСКИЕ СВОЙСТВА И ХИМИЧЕСКИЙ СОСТАВ НОРМАЛЬНОЙ МОЧИ. ПАТОБИОХИМИЯ ПОЧЕК. ПАТОЛОГИЧЕСКИЕ КОМПОНЕНТЫ МОЧИ
ОСОБЕННОСТИ СТРОЕНИЯ ПОЧЕК
Масса обеих почек у взрослого человека составляет около
В ткани почки выделяют два слоя: внешний – корковое вещество и внутренний – мозговое вещество. Основная структурно-функциональная единица почечной паренхимы – нефрон. В обеих почках человека их около 2 млн, у крысы – 62000, у собаки – 816000. В нефроне млекопитающих выделяют почечное (мальпигиево) тельце, состоящее из сосудистого клубочка и двухслойной капсулы клубочка (капсула Боумена) и системы канальцев нефрона.
От капсулы клубочка отходит почечный каналец, который в корковом веществе является проксимальной частью канальца нефрона, переходящей в петлю нефрона (петля Генле). В петле выделяют нисходящую и восходящую части. Последняя переходит в дистальную часть канальца нефрона, впадающего в собирательные почечные трубочки. По нескольку собирательных трубочек впадают в сосочковые протоки, открывающиеся в почечные чашки. В почке млекопитающих различают два типа нефронов: корковые нефроны (85%), почечное тельце которых локализуется в корковом веществе, и юкстамедуллярные нефроны (15%), клубочки которых расположены на границе коркового и мозгового вещества, а петля с нисходящей и входящей частями – в мозговом веществе.
http://www.youtube.com/watch?v=glu0dzK4dbU
МЕХАНИЗМ ОБРАЗОВАНИЯ МОЧИ
В нефроне происходят три главных процесса: фильтрация в клубочках, реабсорбция и секреция в канальцах.
Клубочковая фильтрация. Начальным этапом образвания мочи является фильтрация: в почечном тельце из капиллярного клубочка в полость капсулы фильтруется жидкая часть крови. Клубочковая фильтрация – это пассивный процесс.
http://www.youtube.com/watch?v=lH9IXpp5zTU
В условиях покоя у взрослого человека около 1/4 крови, выбрасываемой в аорту левым желудочком сердца, поступает в почечные артерии. Иными словами, через обе почки у взрослого мужчины проходит около 1300 мл крови в минуту, у женщин несколько меньше. Общая фильтрационная поверхность клубочков почек составляет примерно
В норме объемная скорость фильтрации в среднем составляет 125 мл/мин, что в 100 раз превышает продукцию конечной мочи. Скорость фильтрации обеспечивается фильтрационным давлением, которое можно выразить следующей формулой:
ФД = КД – (ОД + КапсД),
где ФД – фильтрационное давление; КД – капиллярное давление; ОД – онкотическое давление; КапсД – внутрикапсулярное давление. Следовательно, для обеспечения процесса фильтрации необходимо, чтобы гидростатическое давление крови в капиллярах превышало сумму онкотического и внутрикап-сулярного. В норме эта величина составляет около 40 гПа (
Строение юкстамедуллярного (а) и коркового (б) нефронов. I – корковое вещество; II – мозговое вещество; А – наружная зона мозгового вещества; Б – внутренняя зона мозгового вещества; 1 – сосудистый клубочек; 2 – капсула почечного клубочка; 3 – проксимальный каналец (извитая часть); 4 – проксимальный каналец (прямая часть); 5 – нисходящее тонкое колено петли нефрона; 6 – восходящее тонкое колено петли нефрона; 7 – восходящее толстое колено петли нефрона; 8 – дистальный извитой каналец; 9 – связующий каналец; 10 – собирательная трубка; 11 – собирательная почечная трубочка.
http://www.youtube.com/watch?v=aQZaNXNroVY&feature=related
Капиллярное давление в почках зависит не столько от артериального давления, сколько от соотношения просвета «приносящей» и «выносящей» артериол клубочка. «Выносящая» артериола примерно на 30% меньше в диаметре, чем «приносящая», регуляция их просвета осуществляется прежде всего кининовой системой. Сужение «выносящей» артериолы увеличивает фильтрацию. Напротив, сужение «приносящей» артериолы снижает фильтрацию. По величине клубочковой фильтрации судят о фильтрационной способности почек. Если в кровяное русло ввести вещество, которое фильтруется в клубочках, но не реабсорбируется и не секретируется канальцами нефро-нов, то его клиренс численно равен объемной скорости клубочковой фильтрации. Клиренс (очищение) любого соединения принято выражать количеством миллилитров плазмы, которое в 1 мин полностью освобождается от определенного вещества при прохождении ее через почки. Веществами, по которым чаще определяют клубочковую фильтрацию, являются инулин и маннитол. Для расчета клиренса (например, инулина) необходимо величину минутного диуреза умножить на Kм/Kкp(отношение концентраций данного вещества в моче и плазме крови):
где С – клиренс; Км – концентрация данного соединения в моче; Ккр – концентрация в плазме крови; V – количество мочи в 1 мин, мл. Например, при расчете клиренса инулина в норме получим величину клубочковой фильтрации, равную 100–125 мл за 1 мин.
Реабсорбция и секреция. Суточное количество ультрафильтрата в 3 раза превышает общее количество жидкости, содержащейся в организме. Естественно, что первичная моча во время движения по почечным канальцам отдает большую часть своих составных частей, особенно воду, обратно в кровь. Лишь 1 % жидкости, профильтрованной клубочками, превращается в мочу. В канальцах реабсорбируется 99% воды, натрия, хлора, гидрокарбоната, аминокислот, 93% калия, 45% мочевины и т.д.
Из первичной мочи в результате реабсорбции образуется вторичная, или окончательная, моча, которая затем поступает в почечные чашки, лоханку и по мочеточникам попадает в мочевой пузырь. Функциональное значение отдельных почечных канальцев в процессе мочеобразования неодинаково. Клетки проксимального сегмента нефрона реабсорбируют попавшие в фильтрат глюкозу, аминокислоты, витамины, электролиты; 6/7 жидкости, составляющей первичную мочу, подвергается реабсорбции также в проксимальных канальцах. Вода первичной мочи частично (парциально) реабсорбируется в дистальных канальцах. В этих же канальцах происходит дополнительная реабсорбция натрия, могут секрети-роваться в просвет нефрона ионы калия, аммония, водорода и др. В настоящее время в значительной степени изучены молекулярные механизмы реабсорбции и секреции веществ клетками почечных канальцев. Так, установлено, что при реабсорбции натрий пассивно поступает из просвета канальца внутрь клетки, движется по ней к области базальной плазматической мембраны и с помощью «натриевого насоса» поступает во внеклеточную жидкость. До 80% энергии АТФ в клетках канальцев почек расходуется на «натриевый насос». Всасывание воды в проксимальном сегменте происходит пассивно в результате активного всасывания натрия. Вода в этом случае «следует» за натрием. Кстати, в дистальном сегменте всасывание воды происходит вне всякой зависимости от всасывания ионов натрия; этот процесс регулируется антидиуретическим гормоном. Калий в отличие от натрия может не только реабсорбироваться, но и секретироваться. При секреции калий из межклеточной жидкости поступает через базальную плазматическую мембрану в клетку канальца за счет работы «натрий-калиевого насоса», а затем выделяется в просвет нефрона через апикальную клеточную мембрану пассивно. Секреция, как и реабсорб-ция, является активным процессом, связанным с функцией клеток канальцев. Механизмы секреции те же, что и механизмы реабсорбции, но только все процессы протекают в обратном направлении – от крови к канальцу. Вещества, которые не только фильтруются через клубочки, но и реабсор-бируются или секретируются в канальцах, имеют клиренс, который показывает целостную работу почек (смешанный клиренс). В зависимости от того, комбинируется ли фильтрация с реабсорбцией или с секрецией, выделяют два вида смешанного клиренса: фильтрационно-реабсорбционный и фильтрационно-секреционный. Величина смешанного фильтрационно-реабсорбционного клиренса меньше величины клубочкового клиренса, так как часть вещества реабсорбируется из первичной мочи в канальцах. Значение этого показателя тем меньше, чем эффективнее реабсорбция в канальцах.
Так, для глюкозы в норме он равен 0. Максимальное всасывание глюкозы в канальцах составляет 350 мг/мин. Максимальную способность канальцев к обратному всасыванию принято обозначать Тм (транспорт максимум). Иногда встречаются пациенты с заболеванием почек, которые, несмотря на высокое содержание глюкозы в плазме крови, не выделяют глюкозу с мочой, так как фильтруемое количество глюкозы ниже значения Тм. Наоборот, при врожденном заболевании почечная глюкозурия может быть основана на снижении значения Тм.
Регуляция реабсорбции в почке (схема по А.П. Зильберу).
http://www.youtube.com/watch?v=KINOArtDeWg&feature=related
Для мочевины величина смешанного фильтрационно-реабсорбционного клиренса составляет 70. Это значит, что из каждых 125 мл ультрафильтрата или плазмы крови за минуту от мочевины полностью освобождаются 70 мл. Иными словами, определенное количество мочевины, а именно то, которое содержится в 55 мл ультрафильтрата или плазмы, всасывается обратно. Величина смешанного фильтрационно-секреционного клиренса может быть больше клубочкового клиренса, так как к первичной моче прибавляется дополнительное количество вещества, которое секретируется в канальцах. Этот клиренс тем больше, чем сильнее секреция канальцев. Клиренс некоторых веществ, секретируемых канальцами (например, диодраст, пара-аминогиппуровая кислота), настолько высок, что практически приближается к величине почечного кровотока (количество крови, которое за минуту проходит через почки). Таким образом, по клиренсу этих веществ можно определить величину кровотока. Реабсорбция и секреция различных веществ регулируются ЦНС и гормональными факторами. Например, при сильных болевых раздражениях или отрицательных эмоциях может возникнуть анурия (прекращение процесса мочеобразования). Всасывание воды возрастает под влиянием антидиуретического гормона вазопрессина. Альдостерон увеличивает реабсорбцию натрия в канальцах, а вместе с ним и воды. Всасывание кальция и фосфата изменяется под влиянием паратиреоидного гормона. Паратгормон стимулирует секрецию фосфата, а витамин D задерживает ее.
При недостаточном поступлении крови к почечным клубочкам, сопровождающемся небольшим растяжением стенок артериол (снижение давления), происходит возбуждение заложенных в стенках артериол клеток юкстагло-мерулярного аппарата (ЮГА). Они начинают усиленно секретировать протеолитический фермент ренин, катализирующий начальный этап образования ангиотензина.
Субстратом ферментативного действия ренина является ангиотензиноген (гликопротеин), относящийся к α2-глобулинам и содержащийся в плазме крови и лимфе. Ренин разрывает в молекуле ангиотензиногена пептидную связь, образованную двумя остатками лейцина, в результате чего освобождается дека-пептид ангиотензин I, биологическая активность которого незначительна в среде, близкой к нейтральной. Считают, что под влиянием специальной пептидазы, обнаруженной в плазме крови и тканях,– ангиотензин I превращающего фермента (дипеп-тидил-карбоксипептидаза I) из ангиотензина I образуется октапептид ан-гиотензин II. Главным местом этого превращения являются легкие. В
РОЛЬ ПОЧЕК В ПОДДЕРЖАНИИ КИСЛОТНО-ОСНОВНОГО РАВНОВЕСИЯ
Почки оказывают значительное влияние на кислотно-основное равновесие, но оно сказывается по истечении значительно большего времени, чем влияние буферных систем крови и легких. Влияние буферных систем крови обнаруживается в течение 30 с. Легким требуется примерно 1–3 мин, чтобы сгладить наметившийся сдвиг концентрации водородных ионов в крови, почкам необходимо около 10–20 ч для восстановления нарушенного кислотно-основного равновесия.
Основным механизмом поддержания концентрации водородных ионов в организме, реализуемым в клетках почечных канальцев, являются процессы реабсорбции натрия и секреции ионов
водорода. Этот механизм осуществляется с помощью нескольких химических процессов. Первый из них – реабсорбция натрия при превращении двузамещенных фосфатов в однозамещенные. Почечный фильтрат, формирующийся в клубочках, содержит достаточное количество солей, в том числе и фосфатов. Однако концентрация двузамещенных фосфатов постепенно убывает по мере продвижения первичной мочи по почечным канальцам. Так, в крови отношение однозаме-щенного фосфата к двузамещенному составляет 1:4, в клубочковом фильтрате – 9:1, в моче, которая проходит через дистальный сегмент нефрона,– 50:1. Это объясняется избирательным всасыванием канальцевыми клетками ионов натрия. Вместо них из канальцевых клеток в просвет почечного канальца выделяются ионы водорода. Таким образом, двузамещенный фосфат Na2HPO4 превращается в однозамещенный NaH2PO4 и в таком виде выделяется с мочой. В клетках канальцев из угольной кислоты образуется бикарбонат, увеличивая тем самым щелочной резерв крови. Второй химический процесс, который обеспечивает задержку натрия в организме и выведение излишка водородных ионов,– это превращение в просвете канальцев бикарбонатов в угольную кислоту. В клетках канальцев при взаимодействии воды с углекислым газом под влиянием карбоан-гидразы образуется угольная кислота. Водородные ионы угольной кислоты выделяются в просвет канальца и соединяются там с анионами бикарбоната; эквивалентный этим анионам натрий поступает в клетки почечных канальцев. Образовавшаяся в просвете канальца Н2СО3 легко распадается на СО2 и Н2О и в таком виде покидает организм. Третьим процессом, который также способствует сохранению натрия в организме, является образование в почках аммиака, который используется вместо других катионов для нейтрализации и выведения кислых эквивалентов с мочой. Основным источником этого служат процессы дезаминирования глутамина, а также окислительного дезаминирования аминокислот, главным образом глутаминовой кислоты. Распад глутамина происходит при участии фермента глутаминазы, при этом образуются глутаминовая кислота и свободный аммиак. Глутаминаза обнаружена в различных органах и тканях человека, однако наибольшая ее активность отмечена в тканях почек. В общем итоге соотношение концентрации водородных ионов в моче и крови может составить 800:1 – настолько велика способность почек выводить из организма ионы водорода. Процесс усиливается в тех случаях, когда возникает тенденция к накоплению ионов водорода в организме.
НЕКОТОРЫЕ ОСОБЕННОСТИ ОБМЕНА ВЕЩЕСТВ В ПОЧЕЧНОЙ ТКАНИ В НОРМЕ И ПРИ ПАТОЛОГИИ
Сложные физиологические процессы в почечной ткани протекают с постоянным потреблением большого количества энергии, выделяемой при метаболических реакциях. Не менее 8–10% всего поглощаемого человеком в покое кислорода используется на окислительные процессы в почках. Потребление энергии на единицу массы в почках больше, чем в любом другом органе. В корковом веществе почки ярко выражен аэробный тип обмена веществ. В мозговом веществе преобладают анаэробные процессы. Почка относится к органам, наиболее богатым ферментами. Большинство этих ферментов встречается и в других органах. Так, ЛДГ, АсАТ, АлАт, глутаматдегидрогеназа широко представлены как в почках, так и в других тканях. Вместе с тем имеются ферменты, которые в значительной степени специфичны для почечной ткани. К таким ферментам прежде всего относится глицин-амидинотроансфераза (трансамидиназа). Данный фермент содержится в тканях почек и поджелудочной железы и практически отсутствует в других тканях. Глицин-амидинотрансфераза осуществляет перенос амидиновой группы с L-аргинина на глицин с образованием L-орнитина и гликоциамина:
L-аргинин + Глицин —> L-орнитин + Гликоциамин.
Эта реакция является начальным этапом синтеза креатина. Глицин-амидинотрансфераза была открыта еще в
ОБЩИЕ СВОЙСТВА И СОСТАВНЫЕ ЧАСТИ МОЧИ
Общие свойства мочи
Количество выделяемой за сутки мочи (диурез) в норме у взрослых людей колеблется от 1000 до 2000 мл и составляет в среднем 50–80% от объема принятой жидкости. Суточное количество мочи ниже 500 мл и выше 2000 мл у взрослых считается патологическим. Увеличение объема мочи (полиурия) наблюдается при приеме большого количества жидкости, употреблении пищевых веществ, повышающих диурез (арбуз, тыква и др.). При патологии полиурия отмечается при заболеваниях почек (хронические нефриты и пиелонефриты), сахарном диабете и других патологических состояниях. Большое количество мочи выделяется при несахарном диабете (diabetes insipidus) –
http://www.youtube.com/watch?v=6x5pVoMb_vI&feature=related
Нормальная моча прозрачна. Мутность мочи может быть вызвана солями, клеточными элементами, бактериями, слизью, жиром (липурия). Причину помутнения мочи можно определить либо под микроскопом (исследование осадка мочи), либо путем химического анализа. Относительная плотность мочи у взрослого человека в течение суток колеблется в довольно широких пределах (от 1,002 до 1,035), что связано с периодическим приемом пищи, воды и потерей жидкости организмом (потоотделение и др.). Чаще она равна 1,012–1,020. Плотность мочи дает определенное представление о количестве растворенных в ней веществ. В сутки с мочой выделяется от 50 до
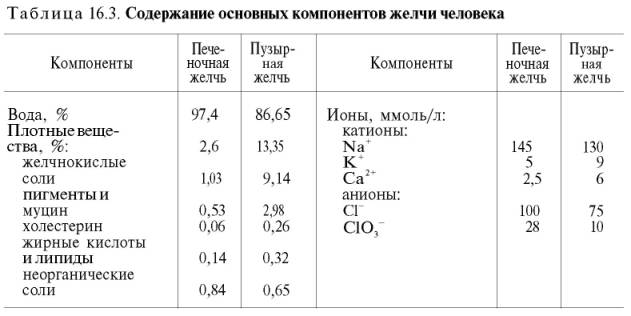
Химический состав мочи
Плотные вещества мочи (около
Всего в моче в настоящее время обнаружено более 150 химических ингредиентов. Далее представлены данные лишь о наиболее важных компонентах мочи человека в норме и при некоторых патологических состояниях.
Органические вещества мочи
Мочевина составляет большую часть органических веществ, входящих в состав мочи. В среднем за сутки с мочой взрослого человека выводится около
Креатинин также является конечным продуктом азотистого обмена. Он образуется в мышечной ткани из фосфокреатина. Суточное выделение креатинина для каждого человека – величина довольно постоянная и отражает в основном его мышечную массу. У мужчин на каждый
Креатин в моче взрослых людей в норме практически отсутствует. Он появляется либо при употреблении значительных количеств креатина с пищей, либо при патологических состояниях. Как только уровень креатина в сыворотке крови достигает 0,12 ммоль/л, он появляется в моче. В первые годы жизни ребенка возможна «физиологическая креатинурия». По-видимому, появление креатина в моче детей раннего возраста обусловлено усиленным синтезом креатина, опережающим развитие мускулатуры. Некоторые исследователи к физиологическим явлениям относят и креатинурию стариков, которая возникает как следствие атрофии мышц и неполного использования образующегося в печени креатина. Наибольшее содержание креатина в моче наблюдается при патологических состояниях мышечной системы и прежде всего при миопатии, или прогрессирующей мышечной дистрофии. Принято считать, что креатин в моче (креатинурия) больных миопатией может появляться в результате нарушения в скелетной мускулатуре процессов его фиксации (удержания) и фосфорилирования. Если нарушен процесс синтеза фосфокреати-на, то не образуется и креатинин; содержание последнего в моче резко снижается.
В результате креатинурии и нарушения синтеза креатинина резко повышается креатиновый показатель мочи: (количество креатина + количество креатинина)/(количество креатинина). В норме этот показатель близок к 1,1. Известно также, что креатинурию можно наблюдать при поражениях печени, сахарном диабете, эндокринных расстройствах (гипертиреоз, адди-сонова болезнь, акромегалия и др.), инфекционных заболеваниях.
Аминокислоты в суточном количестве мочи составляют около
Известны также врожденные болезни: гиперпролинемия (возникает в результате недостатка фермента пролиноксидазы, следствие – пролину-рия); гипервалинемия (врожденное нарушение обмена валина, что сопровождается резким повышением концентрации валина в моче); цитрул-линемия (врожденное нарушение цикла образования мочевины, обусловленное недостатком фермента аргининсукцинат-синтетазы, с мочой выделяется увеличенное количество цитруллина) и др.
Мочевая кислота является конечным продуктом обмена пуриновых оснований. За сутки с мочой выделяется около
Гиппуровая кислота в небольшом количестве всегда определяется в моче человека (около
Безазотистые органические компоненты мочи – это щавелевая, молочная и лимонная (цитрат), а также масляная, валериановая, янтарная (сукцинат), β-оксимасляная, ацетоуксусная и другие кислоты. Общее содержание органических кислот в суточном количестве мочи обычно не превышает
Неорганические (минеральные) компоненты мочи
В моче содержатся практически все минеральные вещества, которые входят в состав крови и других тканей организма. Из 50–65 г сухого остатка, образующегося при выпаривании суточного количества мочи, на долю неорганических компонентов приходится 15–25 г.
Ионы натрия и хлора. В норме около 90% принятых с пищей хлоридов выделяется с мочой (8–15 г NaCl в сутки). При ряде патологических состояний (хронический нефрит, диарея, острый суставной ревматизм и др.) выведение хлоридов с мочой может быть снижено. Максимальная концентрация ионов Na+и Сl– (в моче по 340 ммоль/л) может наблюдаться после введения в организм больших количеств гипертонического раствора.
Ионы калия, кальция и магния. Многие исследователи считают, что практически все количество ионов калия, которое имеется в клубочковом фильтрате, всасывается обратно из первичной мочи в проксимальном сегменте нефрона. В дистальном сегменте происходит секреция ионов калия, которая в основном связана с обменом между ионами калия и водорода. Следовательно, обеднение организма калием сопровождается выделением кислой мочи.
Ионы Са2+ и Mg2+ выводятся через почки в небольшом количестве (см. табл. 18.1). Принято считать, что с мочой выделяется лишь около 30% всего количества ионов Са2+ и Mg2+, подлежащего удалению из организма. Основная масса щелочноземельных металлов выводится с калом.
Бикарбонаты, фосфаты и сульфаты. Количество бикарбонатов в моче в значительной мере коррелирует с величиной рН мочи. При рН 5,6 с мочой выделяется 0,5 ммоль/л, при рН 6,6 – 6 ммоль/л, при рН 7,8 – 9,3 ммоль/л бикарбонатов. Уровень бикарбонатов повышается при алкалозе и понижается при ацидозе. Обычно с мочой выводится менее 50% всего количества выделяемых организмом фосфатов. При ацидозе выведение фосфатов с мочой возрастает. Повышается содержание фосфатов в моче при гиперфункции паращитовидных желез. Введение в организм витамина D снижает выделение фосфатов с мочой.
Серосодержащие аминокислоты: цистеин, цистин и метионин – являются источниками сульфатов мочи. Эти аминокислоты окисляются в тканях организма с образованием ионов серной кислоты. Общее содержание сульфатов в суточном количестве мочи обычно не превышает
Аммиак. Как отмечалось, существует специальный механизм образования аммиака из глутамина при участии фермента глутаминазы, которая в большом количестве содержится в почках. Аммиак выводится с мочой в виде аммонийных солей. Содержание последних в моче человека в определенной степени отражает кислотно-основное равновесие. При ацидозе их количество в моче увеличивается, а при алкалозе снижается. Содержание аммонийных солей в моче может быть снижено при нарушении в почках процессов образования аммиака из глутамина.
Патологические компоненты мочи
Широко используемое понятие «патологические компоненты мочи» в известной мере условно, так как большинство соединений, рассматриваемых как патологические компоненты мочи, хотя и в небольшом количестве, но всегда присутствуют в нормальной моче. Иными словами, речь идет о веществах, которые в нормальной моче не встречаются в аналитически определяемых количествах. Это прежде всего белки, глюкоза, ацетоновые (кетоновые) тела, желчные и кровяные пигменты.
Белок. В нормальной моче человека содержится минимальное количество белка, присутствие которого не может быть доказано обыкновенными качественными пробами на наличие белка. При ряде заболеваний, особенно при болезнях почек, содержание белка в моче может резко возрасти (протеинурия). Источником белка мочи являются белки сыворотки крови, а также в какой-то степени белки почечной ткани.
Протеинурии делятся на две большие группы: почечные и внепочечные. При почечных протеинуриях белки (в основном белки плазмы крови) попадают в мочу вследствие органического повреждения нефрона, увеличения размеров пор почечного фильтра, а также в результате замедления тока крови в клубочках. Внепочечные протеинурии обусловлены поражением мочевых путей или предстательной железы.
Часто употребляемое в клинической практике название «альбуминурия» (при обнаружении в моче белка) неправильно, так как с мочой выделяются не только альбумины, но и глобулины. Например, при нефрозах общее содержание белка в моче может достигать 26 г/л, при этом концентрация альбуминов 12 г/л, аглобулинов – 14 г/л. В моче человека можно обнаружить активность ряда ферментов: липазы, рибонуклеазы, ЛДГ, аминотрансфераз, урокиназы, фосфатаз, α-амила-зы, лейцинаминопептидазы и др. Основные трудности при определении активности ферментов мочи, кроме α-амилазы и некоторых других, заключаются в необходимости сгущения (концентрирования) мочи и предотвращении ингибирования ферментов в процессе этого сгущения.
Кровь. В моче кровь может быть обнаружена либо в форме красных кровяных клеток (гематурия), либо в виде растворенного кровяного пигмента (гемоглобинурия). Гематурии бывают почечные и внепочеч-ные. Почечная гематурия – основной симптом острого нефрита. Внепочеч-ная гематурия наблюдается при воспалительных процессах или травмах мочевых путей. Гемоглобинурии обычно связаны с гемолизом и гемоглоби-немией. Принято считать, что гемоглобин появляется в моче после того, как содержание его в плазме превысит
Глюкоза. Нормальная моча человека содержит минимальные количества глюкозы, которые не обнаруживаются обычными качественными пробами. При патологических состояниях содержание глюкозы в моче увеличивается (глюкозурия). Например, при сахарном диабете количество глюкозы, выделяемое с мочой, может достигать нескольких десятков граммов в сутки.
Иногда в моче обнаруживают и другие углеводы, в частности фруктозу, галактозу, пентозу. Фруктозурия наблюдается при врожденной недостаточности ферментов, превращающих фруктозу в глюкозу; встречаются также и врожденная пентозурия, и врожденная галактозурия.
Кетоновые (ацетоновые) тела. В нормальной моче эти соединения встречаются лишь в самых ничтожных количествах (не более
Билирубин. В норме моча содержит минимальное количество билирубина, которое не может быть обнаружено обычными качественными пробами. Повышенное выделение билирубина, при котором обычные качественные пробы на наличие билирубина в моче становятся положительными, называется билирубинурией. Она встречается при закупорке желчного протока и заболевании паренхимы печени. Выделение билирубина в мочу особенно сильно выражено при обтура-ционных желтухах. При застое желчи переполненные желчью канальцы травмируются и пропускают билирубин в кровяные капилляры. Если поражена паренхима печени, билирубин проникает в кровь через разрушенные печеночные клетки. Билирубинурия проявляется при уровне прямого билирубина в крови выше 3,4 мкмоль/л. Непрямой билирубин не может пройти через почечный фильтр. Это становится возможным при значительных поражениях почек.
Уробилин. В моче уробилин, точнее стеркобилин, присутствует всегда в незначительном количестве. Концентрация его резко возрастает при гемолитической и печеночной желтухах. Это связано с потерей печенью способности задерживать и разрушать мезобилиноген (уробилиноген), всосавшийся из кишечника. Напротив, отсутствие в моче уробилиногена при наличии желчных пигментов (билирубина) указывает на прекращение поступления желчи в кишечник вследствие закупорки желчного протока.
Порфирины. В норме моча содержит лишь очень малые количества порфиринов I типа (до 300 мкг в суточном количестве). Однако выделение порфиринов может резко возрасти (в 10–12 раз) при заболеваниях печени и пернициозной анемии. При врожденной порфирии имеет место сверхпродукция порфиринов I типа (уропорфирина I и копропорфирина I). В этих случаях в суточном количестве мочи обнаруживается до 10 мг смеси этих порфиринов. При острой порфирии отмечается экскреция с мочой повышенных количеств уропорфирина III, копропорфирина III, а также порфобилиногена.
Мочевые камни
Мочевые камни – это плотные образования, встречающиеся в мочевыводя-щих путях. Мочевые камни могут располагаться в паренхиме почек, в чашках, лоханках, мочеточниках, мочевом пузыре и мочеиспускательном канале. Величина, форма и консистенция мочевых камней разнообразны. Мелкие мочевые камни имеют вид песчинок, большое количество которых образует так называемый мочевой песок. Более крупные мочевые камни обычно имеют округлую, овальную или, реже, корраловидную форму. Общим в структуре мочевых камней является наличие так называемого ядра, вокруг которого расположена различной толщины оболочка, или тело камня. Примерно треть или более таких камней состоит из Са3(РО4)2, MgNH4PO4, CaC2O4или их смесей, т.е. это щавелевокислые (оксалатные), фосфорнокислые (фосфатные) или смешанные мочевые камни. Часто образование камней происходит в результате хронического защелачивания мочи в мочевом пузыре и почечных лоханках, которое является следствием бактериальной инфекции. Образованию камней способствуют избыточное выделение ионов Са2+, например, при гиперпаратиреоидозе, остеопорозе (в частности, вызванном неподвижностью) и необычайно высокое содержание Са2+ в пище. Кроме того, камни, состоящие из оксалата кальция, пато-гномоничны для оксалурии (наследственное нарушение метаболизма глицина, при котором практически весь синтезированный глицин окисляется через глиоксиловую кислоту до щавелевой кислоты). У больных подагрой, как правило, встречаются камни, состоящие в основном из мочевой кислоты (C5H4N4O3), реже – из ее аммониевой или натриевой соли. Эти камни получили название мочекислых, или уратных. Отложение цистина (цистиновые камни) почти постоянно наблюдается у больных цистинурией. Следует отметить, что изучение этиологических факторов, определение химического состава мочевых камней имеют важное значение для профилактики и лечения почечнокаменной болезни.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}