Тема 2.
Клітинні включення рослин (запасні та екскреторні речовини). Оболонка рослинної клітини.
Клітини рослин є дуже різноманітними і своєрідними: від багатоклітинних нитчастих водоростей і одноклітинних рослин до судинних рослин. У хлорели одній клітині доводиться виконувати декілька функцій, які у багатоклітинних рослин поділені між клітинами різних тканин. В той же час навіть клітини, що дуже відрізняються між собою, володіють глибокою схожістю в будові і функціях.
![]()
![]()
![]()

Це зумовлене тим, що всі клітини організму (якщо виключити вегетативне розмноження, при якому від материнської рослини відразу відділяється як нова особина особа або її зачаток цілий багатоклітинний комплекс) є нащадками однієї і тієї ж клітини-родоначальниці. Тому, яким би чином не були спеціалізовані клітини, вони мають загальне походження.
Загальні риси межі в будові клітин рослин різних видів пов’язані з тим, що всі рослини в тому або іншому ступені споріднені. Всі рослини розвинулися шляхом еволюції від одноклітинних спільних предків. Загальні риси межі успадковані клітками сучасних рослин від стародавніх клітин. З цим зв’язана також наявність загальних рис меж будови та роботи рослинних і тваринних клітин.
Склад клітини |
||
|
Складова |
Будова |
Функції |
|
Целюлоза і пектини, у тваринної клітини ззовні глікокалікс |
|
|
|
Двомембранна структура, будова – біліпідний шар (що складається з фосфоліпідів, гліколіпідів та холестеролу) та мозаїчно вкраплені інтегральні білки; ззовні – мозаїчно вкраплені вуглеводні (властива напівпроникність) |
|
|
|
Гіалоплазма, органели (ЕПС, мітохондрії, рибосоми тощо) та цитоплазматичні включення |
|
|
|
Рідке середовище (колоїдний розчин) цитоплазми, у якому містяться органели і включення |
|
|
|
Найважливіший немембранний органоїди живої клітини сферичної або злегка еліпсоїдні форми, діаметром 100-200 ангстрем, що складається з великої та малої субодиниць |
|
|
|
Ультрамікроскопічна система мембран, що утворюють трубочки, канальні, цистерни і пухирці; сітка об’єднана із зовнішньою мембраною ядерної оболонки та зовнішньою клітинною мембраною; є два типи – гранулярний (на поверхні – рибосоми) і гладкий (рибосоми на поверхні відсутні) |
|
|
|
Система плоских порожнистих міхурців з трубочками по краях (неактивними лізосомами); у рослинних клітинах містить полісахариди, які використовуються для будови целюлозної оболонки |
|
|
|
Круглясті або овальні утвори з окремою мембраною, що містять більше як 30 ферментів. Формуються з пухирців (везикул), які відділяються від комплекса Гольджі, та ендосом, у які потрапляють речовини при ендоцитозі. Розрізняють первинні (які містять неактивні ферменти) та вторинні (ферменти яких активні) лізосоми. Кількість визначається життєдіяльністю клітини та її фізіологічним станом |
|
|
|
Великі пухирці із переважно водним змістом (мінеральні солі, цукор, пігменти, органічні кислоти, ферменти). Утворюються з пухирців ЕПС або пухирців Гольджі. Розрізняють чотири типи вакуолей – скоротливі, або пульсуючі (осмотична регуляція у прісноводних найпростіших), травні (насамперед у тваринній клітині), фагоцитарні та центральну вакуолю рослинної клітини (виникає з маленьких вакуоль і пухирців ЕПС). Вміст – клітинний сік |
|
|
|
Подвійна мембранна оболонка, внутрішня частина якої утворює вирости – кристи, а зовнішня містить отвори, що складаються зі спеціальних білків. Ці утвори пропускають невеликі молекули та йони. Внутрішня мембрана таких виростів не має – вона складається з молекул АТФ-синтази, які, при проходженні через них протонів, синтезують АТФ. Внутрішня порожнина заповнена матриксом, де містяться ферменти та мітохондріальні ДНК і РНК. Мітохондрія також містить рибосоми, білки, вітаміни і ліпіди. У клітині міститься від 1 до 1500 мітохондрій |
|
|
|
Продукти життєдіяльності клітини, що нагромаджуються в цитоплазмі (крохмаль, глікоген, кристали солей) |
|
|
|
Оболонка ядра (з ядерними порами) і ядерце. Оболонка складається з двох шарів (внутрішнього і зовнішнього, який переходить у ЕПС. У ядрі містяться хромосоми, які, спочатку маючи двохроматидну будову, після поділу ядра стають однохроматидними. Внутрішнє середовище заповнене ядерним соком, до складу якого входять білки-ферменти, необхідні для синтезу нуклеїнових кислот, фібрилярних білків, вуглеводів, мінеральних солей і води. Усередині ядра розміщене ядерце – специфічне тільце, що не має мембрани (у ядрі від 2 до 7 тілець). Складається з білка та РНК. Формуються на певних ділянках окремих хромосом. Під час поділу клітин розпадається. Місце синтезу субодиниць рибосом) |
|
|
Схожість також зумовлена тим, що всі живі клітини, яку б спеціальну функцію вони не несли в організмі, перш за все повинні забезпечувати власне життя. Клітини поглинають поживні речовини, переробляють їх, здобуваючи енергію і будуючи власне тіло, дихають, звільняються від непотрібних речовин, борються з різними шкідниками, реагують на зміни зовнішніх умов, перебудовувавши свою життєдіяльність, ростуть.
Всі ці процеси у різних клітин здійснюються подібно і за допомогою однотипних структур, загальних за планом будови не тільки для різних рослинних клітин, але і для клітин рослин і тварин. Треба сказати, що виконання будь-якої спеціальної функції клітки ґрунтується на її загальних властивостях. Та або інша властивість спеціалізованої клітини клітини розвивається особливо сильно і забезпечує виконання клітиною її основної, спеціальної функції. Мертві спеціалізовані клітини – це крайній приклад цього. Спеціальна функція таких клітин пов’язана з їх оболонкою; протопласт потрібний лише до тих пір, поки він створює оболонку; після цього він відмирає, і вся клітина складається тільки з неживої оболонки, яка і працює на потреби рослини.
Зовні рослинна клітина вкрита оболонкою, неоднаковою по товщині і будові у різних клітин. Її речовини виробляються в цитоплазмі і відкладаються зовні від неї, поступово створюючи оболонку. Цими речовинами перш за все є полісахариди пектин, геміцелюлоза і в невеликих кількостях целюлоза. Вони утворюють так звану первинну оболонку. Вона досить еластична, у міру зростання клітини розтягується і теж росте зростає , а тому не перешкоджає росту клітини. Проте вона створює певну міцність клітини і здатна захистити її від механічних пошкоджень. Є клітини, які позбавлені такої первинної оболонки – зооспори і гамети водоростей та нижчих грибів, чоловічі гамети вищих рослин.
Оболонка — це зовнішній пограничний шар клітини. Це продукт життєдіяльності цитоплазми. У підтвердження цього говорить той факт, що голі клітини, а також зооспори на ранніх етапах свого розвитку не мають оболонки, а пізніше через певний час вкриваються оболонкою. Більшість представників рослинного світу вкриті оболонкою. Вона відсутня лише у статевих клітин (гамет), зооспор, а також у вегетативних клітин деяких нижчих рослин, зокрема у джгутикових, грибів (міксоміцетів, архіміцетів), водоростей та ін. Проте у цих рослин їх протопласт відмежований від зовнішнього середовища еластичним шаром плазматичною мембраною (плазмалемою).
Оболонки рослинних клітин на відміну від тваринних твердіші і вони, в основному, зумовлюють форму клітини. Клітинна оболонка досить міцна і зберігається на протязі всього їх життя. Оболонки деяких клітин навіть після їх відмирання ще деякий час можуть брати участь в життєвих процесах рослинних організмів. Характер клітинної оболонки залежить від функції клітини. Так, наприклад, живі клітини, в яких протікають обмінні процеси, мають тонку оболонку, а клітини, які виконують механічні та провідні функції, мають потовщені оболонки. Морфологія клітинної оболонки може змінюватися в залежності від віку клітин, а також від функцій, які вона виконує. Оболонка, як провило, без кольору і є прозорою, легко пропускає сонячне світло. По ній можуть переміщатися вода розчинені низькомолекулярні речовини. У кожній клітині є власна оболонка. Оболонка сусідніх клітин з’єднані між собою речовинами, які формують так звану серединну пластинку.
2. Молекулярна будова і механізм утворення компонентів оболонки.
Клітинна оболонка побудована , в основному з полісахаридів, мономерами яких є цукри — з’єднані між собою глікозидним зв’язком (-О-) у вигляді ланцюга. Крім моносахаридів в склад клітинної оболонки входять білки, мінеральні солі, лігнін, пігменти, ліпіди, а також вода. Полісахариди поділять на: скелетні речовини і речовини матриксу. Скелетною речовиною оболонки вищих рослин є целюлоза (клітковина), яка в хімічному відношенні представляє собою β-1,4-D-глюкан. Кількість глюкозних залишків в молекулі клітинних оболонок різних рослин різна, що визначає властивості целюлози. Целюлоза досить міцна і еластична, вона не вступає в хімічні реакції, не розчиняється у кислотах. Тільки концентровані соляна і сірчана кислоти руйнують целюлозу, перетворюючи її в цукор. Реактивом для виявлення целюлози є хлор-цинк-йод. Внаслідок взаємодії целюлози з реактивом вона забарвлюється в синьо-фіолетовий колір. У первинній оболонці міститься біля 5 %, у вторинній 85 % целюлози. Полісахариди матриксу представлені пектиновими речовинами і геміцелюлозою. Поширеним представником пектинових речовин є α-1,4-полігалактуронова кислота, або пектинова кислота. Пектинові речовини набухають у воді, деякі розчиняються у ній. Легко руйнуються під дією кислот.
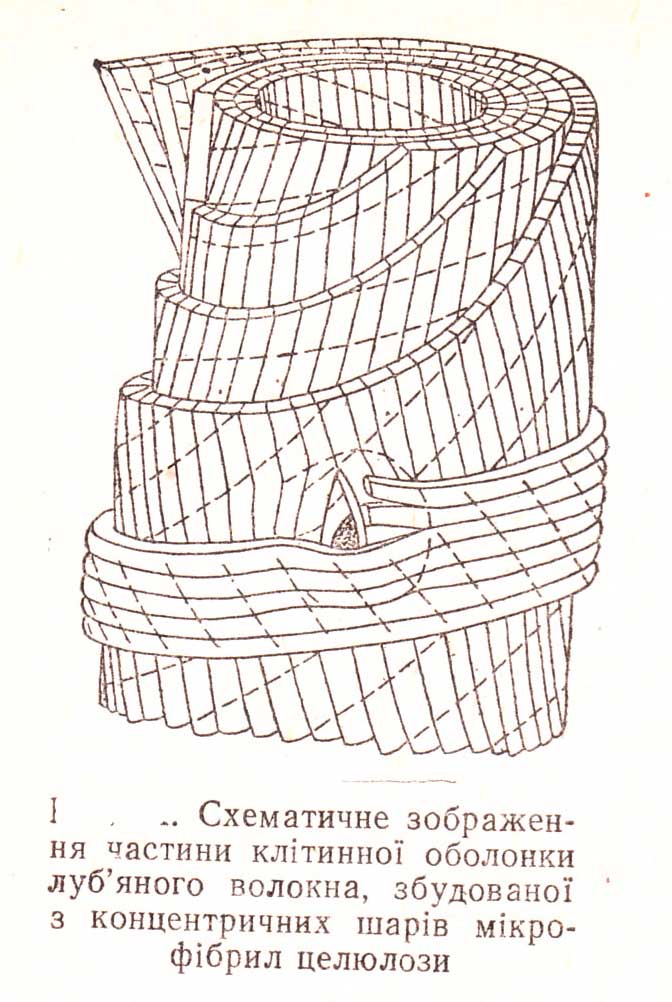
За допомогою електронного мікроскопу досліджено ультраструктуру клітинної оболонки. Встановлено, що полімерна молекула целюлози складається із залишків глюкози, які з’єднані між собою у вигляді ланцюжків. Ланцюжки розміщуються паралельно один до одного, що зумовлює впорядкованість оболонки клітини. З’єднані між собою по декілька десятків (40-60), ланцюжки молекул утворюють елементарні міцели діаметром 50-70Αْ і довжиною біля 600 А˚. Близько розташовані елементарні міцели утворюють мікрофібрили, які представляють собою основну структурну одиницю целюлози. Мікрофібрили (від грец. micros — малий і лат. fibril — волоконце) — основні структурні одиниці целюлози, що являють собою сукупність близько розташованих елементарних міцел. Мікрофібрили розташовуються одна від одної на певній відстані, а проміжки між ними заповнюються пектиновими речовинами. Довжина М. коливається в межах кількох тисяч А˚, а діаметр — від 100 до 250 А˚. Сукупність мікрофібрил утворює вищу структурну одиницю целюлози — фібрили. Фібрили — це пластинки або волокна довжиною декілька мікронів, а діаметром до 0,5 ммк.
Необхідно відмітити, що структура оболонки клітини більш менш постійна для кожного виду рослин і відображає його біологічну специфіку. Однак, у різних рослин структура целюлози може бути досить різноманітною. Хімічна і структурна різноманітність целюлози визначається:
· Числом ланцюжків (ниток) молекул, з яких складаються міцели;
· Числом елементарних міцел, з яких складаються мікрофібрили;
· Числом мікрофібрил, з яких формуються фібрили — пластинки і волокна целюлози.
·
В залежності від хімічної і структурної різноманітності целюлози визначається ї використання в різноманітних галузях промисловості. Важливе значення в структурній будові має просторове розташування міцел. Розрізняють такі способи розташування міцел:
· Міцели розташовуються паралельно до повздовжньої осі клітини (виражена волокниста текстура);
· Міцели розташовуються перпендикулярно до повздовжньої осі (виражена кільчаста структура);
· Міцели розташовуються спірально вздовж повздовжньої осі.
Всі речовини клітинної оболонки синтезуються протопластом самої клітини. Оскільки оболонка продукт життєдіяльності протопласту, то вона завжди відокремлена від нього плазмалемою. У зв’язку з цим речовини оболонки, як правило, виділяються в неї з боку протопласта. Основну роль в утворенні оболонки відіграють апарат Гольджі і плазмалема. Ці мембранні системи містять форменні елементи, які приймають участь в біосинтезі полісахаридів. Синтез полісахаридів матриксу оболонки (з цукрів) починається ще в пухирцях Гольджі після їх відокремлення від країв цистерн. Пухирці Гольджі рухаються до поверхні протопласту і підходять до плазмолеми. Відтак, обмежуюча їх мембрана вбудовується в плазмалему і вміст з полісахаридами виливається в оболонку. У період інтенсивного відкладання речовин в оболонку, плазмалема займає контур.
Апарат Гольджі є місцем синтезу і транспорту лігніну, а також глюкопротеїду оболонки. Синтез і кристалізація целюлози, утворення і орієнтація мікрофібрил, як правило, здійснюється плазмалемою, а необхідні для цього ферменти і, можливо, попередники транспортуються до неї пухирцями Гольджі. Утворювані плазмалемою мікрофібрили накладаються одна на одну зсередини. В орієнтації мікрофібрили велика роль належить мікро трубочкам, оскільки після їх штучного руйнування утворюються лише ізодіаметричну клітини.
3. Структурна організація оболонки.
Початок формування оболонки пов’язаний з процесом поділу клітини. Між дочірніми ядрами материнської клітини в екваторіальній її частині можна спостерігати цитоплазматичну структуру, яка називається фрагмопластом.
Фрагмопласт (від грец. phragmos — загорода i plastos — виліплений, утворений) — зачаткова пластинка клітини, що виникає в екваторіальній площині в телофазі мітозу і бере участь в утворенні клітинної оболонки.
Фрагмопласт закладається ще в анафазі у вигляді невеликих пухирців, що розташовуються впоперек ахромати нового веретена між хромосомами , що відійшли до протилежних полюсів. Пізніше пухирці зливаються і утворюють тонку смужку із напіврідкої цитоплазматичної маси. Таким чином, клітинна оболонка утворюється в протопласті і є продуктом його життєдіяльності. Електромікроскопічними дослідженнями було встановлено, що в утворенні первинної оболонки, яка є продовженням розвитку фрагмопласт, безпосередню участь бера комплекс Гольджі, який направляє до фрагмопласта пухирці, що зливаються з ним. Спочатку серединна пластинка закладається в центрі материнської клітини, а пізніше наростає у відцентровому напрямку до тих пір, доки не зімкнеться з боковими її стінками. Цей процес завершується тоді, коли сформовані дочірні ядра розійшлися до полюсів. Таким чином, серединна пластинка ділить протопласт материнської клітини на дві частини, але для обох дочірніх клітин вона є спеціальною і ніби втримує їх в одному комплексі. Роз’єднані протопласти дочірніх клітин відкладають з обох сторін на серединну пластинку первинну оболонку. В цей час кожна із дочірніх клітин збільшується в розмірах у декілька раз. Первинна стінка відносно тонка, середньому від 0,5 до 1 мк. Під мікроскопом видно, що вона представляє собою гомогенну структуру. Ріст первинної оболонки клітини відбувається шляхом інтусусцепції — впровадження мікрофібрил целюлози в міжмолекулярні їх простори. Однак механізм синтезу мікрофібрил целюлози і відкладання їх в первинній оболонці остаточно ще нез’ясоване. Первинною оболонкою в основному, вкриті клітини меристематичних клітин. Згодом в процесі диференціації клітини на внутрішню сторону первинної стінки (оболонки) накладається вторинна оболонка. Відкладання речовин первинної оболонки забезпечує її ріст в дожину, а вторинної — в товщину. Причому як первинна, так і вторинна оболонки ростуть за рахунок діяльності протопласту. Різні групи клітин різні групи клітин в залежності від спеціалізації мають вторинні оболонки різної товщини. Вторинна оболонка (стінка) наростає шляхом аппозиції, тобто накладання на старі шари міцел внутрішньої поверхні стінки нових. Це призводить до зменшення порожнини клітини. Накладання нових шарів на зовнішню сторону стінки спостерігається досить рідко, наприклад, на поверхню клітин епідерми, спор, пилку. Таким чином, на основі викладеного матеріалу можна представити схему будови оболонки клітини наступним чином: первинна оболонка, вторинна оболонка, третинна оболонка.
4. Первинна оболонка: визначення та утворення.
Плазмодесми, пори і порові поля, як структурні компоненти первинної оболонки.
Первинна оболонка — зовнішній шар, який безпосередньо примикає до міжклітинної речовини. Взаємозв’язок контактуючих клітин здійснюється за допомогою особливих утворень в клітині: пор, перфорацій, плазмодесм.
Пори (від грец. poros — отвір) — мікроскопічно малі не потовщені місця у вторинних оболонках, через які здійснюється обмін між сусідніми клітинами. Розрізняють П.: прості та облямовані.
Проста пора складається з порового каналу, що пронизує вторинну оболонку та замикається первинною. Порові канали сусідніх клітин співпадають і мають однаковий діаметр. Інколи в дуже потовщених оболонках поровий канал розгалужується, внаслідок чого утворюються так звані розгалужені пори, які є в оболонці склереїдів плодів грушу, айви.
Ділянка первинної оболонки, яка міститься в порожнині пори, називається замикаючою пластинкою (плівкою). Через цю плівку речовини з однієї клітини переходять в другу. Таким чином, проста пора складається з каналу пори, що пронизує оболонку та замикається первинною. Прості пори широко поширені і зустрічаються в оболонках клітин різноманітних тканин. Наприклад, в прозенхімних механічних тканинах (волокна льону) зустрічаються косі пори. Вони скоро дегенерують і тому зв’язку між клітинами не виконують.
Порові поля (від грец. poros — отвір) – заглибини у первинній оболонці, що замикаються серединною пластинкою. П.п. сусідніх клітин здебільшого не збігаються.
До складних пор належать облямовані пори. Вони найчастіше утворюються в провідних елементах голонасінних і регулюють водопостачання. Облямована пора складається з порового каналу та камери, тобто лійкоподібної розширеної частини. У сосни, ялини утворюється торус, який представляє собою ділянку серединної пластинки, вкритої з обох боків первинною оболонкою. Діаметр торуса більший. Ніж діаметр отвору пори. Тому, розглядаючи облямовану пору з поверхні клітини вона виглядає як два кола, вписані одне в одне. Менше коло — це канал пори, а більше — куполоподібне підвищення вторинної оболонки, яке оточує канал.
В покритонасінних рослин в стінках водопровідних клітин досить часто виникають отвори, які називають перфораціями.
Перфорації (від лат.perforatio — просверлення, проникнення ) — суцільні отвори в оболонках рослинних клітин. Вони виникають внаслідок розчинення ферментами первинної оболонки і серединної пластинки. П. Утворюються між члениками судин, в перетинках між клітинами ситовидних трубок тощо.
Плазмодесми (від грец. — виліплене, оформлене і — зв’язок, зв’язка) — тонкі цитоплазматичні тяжі , гот проходять через наскрізні отвори в клітинних оболонках і з’єднують протопласти сусідніх клітин у рослин. П. Складається з плазмодесмового каналу, плазмалеми, що вистилає плазмодесмовий канал, десмотрубки, стінки якої неперервні з елементами ендоплазматичного ретикулюму сусідніх клітин та гіалоплазми, що міститься між десмотрубкою і плазмалемою.
За допомогою П. Протопласти клітин в рослинному організмі об’єднуються в єдину систему — симпласт. П. Вперше описав російський ботанік І. Горожанкін (1881).втни властиві лише рослинним клітинам. Утворюються при виникненні клітинних оболонок під час цитокінезу. Внаслідок того, що між пектиновими пухирцями Гольджі, які зливаються між собою залишаються ретикулярні в яких цитоплазма не розходиться між дочірніми клітинами. Стінки канала плазмодесми вистелені плазмалемою, яка формується при злитті пухирців Гольджі. Плазмалема, яка вистеляє канал плазмодесм з плазмалемою суміжних клітин. В центрі плазмодесм проходить трубка (десмотрубки), стінки якої неперервні з мембранами елементів ендоплазматичного ретикулюму обох клітин. Між десмотрубкою і плазмалемою в плазмадесмовому каналі знаходиться гіалоплазма, яка втратила рибосоми і з’єднана з гіалоплазмою суміжних клітин таким чином, дві суміжні клітини зв’язані двома каналами — по гіалоплазмі і по ретикулярному елементу з неперервними, обмежуючими їх мембранами (плазмалемою і мембранами ретикулюму).
5. Вторинна і третинна оболонки: визначення та утворення. Вікові зміни хімічного складу оболонки рослинної клітини: кутинізація, здерев’яніння, опробковіння, ослизнення, мінералізація.
Вторинна оболонка – оболонка клітини, що утворилася внаслідок накладання зсередини на первинну оболонку шарів фібрил целюлози, продукованої протопластом. В.о. утворюється шляхом аппозиції, і нею вкриті клітини епідерми, механічних, провідних та ін. тканин.
Третинна оболонка — внутрішній шар, який контактує з цитоплазматичною мембраною — плазмалемою.
В процесі життєдіяльності рослинних клітин їх оболонки зазнають вікових змін та хімічного складу, зокрема:
Кутинізація ( від лат. cutis — шкірка) — процес відкладання кутину на зовнішніх стінках клітин епідерми.
Здерев’яніння (лігніфікація) (від лат. ligninum — дерево і facio – робити) — процес просочування оболонок рослинних клітин органічною речовиною — лігніном, що надає міцності, стійкості і твердості рослинним тканинам (спостерігається у клітинах вторинної ксилеми).
Опробковіння — процес просочування оболонок рослинних клітин суберином, що призводить до відмирання їх живого вмісту (спостерігається наприклад, при утворенні корку тощо).
Ослизнення — процес видозмінення пектинових речовин і целюлози оболонки у слиз. У водному середовищі слизи дуже набухають, а відтак розріджуються (спостерігається, наприклад, у насінній шкірці льону, гарбуза тощо).
Мінералізація — процес просочування рослинної оболонки мінеральними речовинами. М. обумовлена відкладанням в оболонці клітини мінеральних солей: кремнезему, вуглекислого кальцію та ін. Завдяки мінералізації клітинні оболонки стають дуже твердими.
Апопласт (від грец. apo— без і plastos — утворений ) — система взаємозв’язаних міжфібрилярних просторів клітинних оболонок та міжклітинників, через яку здійснюється переміщення води в рослині.
Симпласт (від грец. syn — разом і plastos — виліплений, утворений) — багатоядерний цитоплазматичний утвір. У рослин С. може мати кілька значень: 1) багатоядерний протопласт, що не має клітинної будови (напр., у водорості каулерпи); 2) протоплазматичний вміст кількох або багатьох злитих клітин (напр., вміст членистих молочників, периплазмодій); 3) сукупність протопластів, з’єднаних плазмодесмами.
Оболонка рослинних клітин має важливе значення. Вона надає рослинним клітинам форму і міцності. Крім того. Вона захищає протопласт від фізичного та механічного впливу, від мікроорганізмів та бактеріальних хвороб. Клітинна оболонка регулює надходження речовин із зовнішнього середовища в клітину, а також їх пересування з одної клітини в іншу. В процесі еволюції рослинного світу оболонка сформувалась в міцну структуру, оскільки в рослин відсутні активні рухи, тому клітини повинні мати надійний захист від несприятливих факторів навколишнього середовища.
Рослинні оболонки широко використовуються в різних галузях народного господарства. Сировиною для деревообробної, паперової, текстильної промисловості є рослинні оболонки. Високосортний папір виробляється з деревини швидкорослих порід (тополя, верба), в яких деревні волокна досягають декількох міліметрів завдовжки. При відповідній обробці з целюлози виготовляють вибухові речовини. Деревина, а значить і оболонки клітин використовується для виробництва спирту, штучного шовку (віскози), для одержання смол, фарб, лаків. Деревина є незамінним матеріалом для виробництва меблів, для деревообробної промисловості. Застосовується і у будівництві. Луб’яні волокна льону, конопель — це цінний матеріал для виробництва тканин. Оболонки рослинних клітин використовуються для виготовлення медикаментів тощо.
Луб’яні волокна льону
Чистими целюлозними стінками є вата і бинт
Клітини, що мають вторинну оболонку, досить міцні. Вони утворюють механічні, опорні тканини рослини. Іноді вторинна оболонка грає і роль складу поживних речовин.
У процесі остаточної диференціації рослинні клітини формують різноманітні скульптурні потовщення своєї оболонки. Розрізняють:
1. Зовнішні потовщення, що бувають на поверхні оболонки клітини і які не з’єднані або частково з’єднані з іншими клітинами. Вони зустрічаються на оболонці пилкових зерен у вигляді гачечків, шипиків, сіточки та на зовнішніх стінках клітин епідерми.
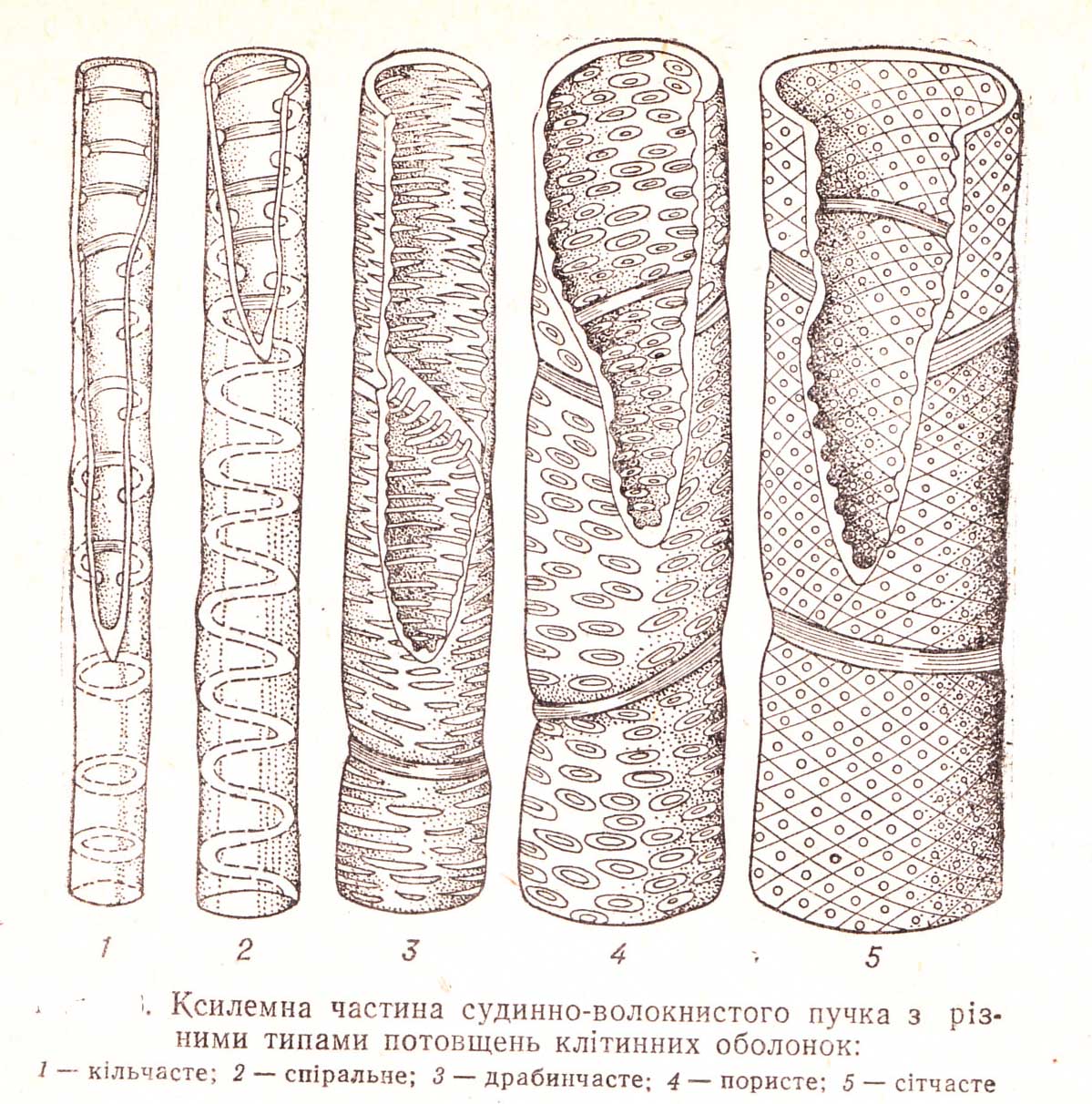
2. Внутрішні потовщення, що утворюють потовщення різного характеру внаслідок нерівномірного відкладання вторинних шарів. Вони бувають у вигляді спіралі різної крутизни і частоти витків, подвійної спіралі, у вигляді кілець різної масивності. Залежно від цього розрізняють такі типи потовщень як кільчасті, спіральні, сітчасті, драбинчасті. Існують також проміжні типи потовщень: кільчасто-спіральне, спірально-кільчасте…
Спіральні, кільчасті, сітчасті та пористі потовщення характерні для ксилемних елементів – судин, що утворюються в результаті злиття поздовжніх рядів клітин. По таких судинах вода рухається з однієї ділянки рослини в іншу, а також просочується крізь пори стінок для постачання сусідніх клітин.
Різні типи потовщення клітинної оболонки відповідають певним етапам онтогенезу рослини. Спіральні та кільчасті судини характерні для ранніх фаз розвитку, коли орган ще росте. Дуже потовщені судини під час росту перешкоджали б розтягуванню органа в довжину, а кільчасті і спіральні потовщення не протидіють його видовженню. Коли ріст органа в довжину закінчується, кільчасті і спіральні судини доповнюються значно міцнішими сітчастими, драбинчастими і пористими.
|
Особливим типом потовщення клітинної оболонки є утворення цистолітів – своєрідних гроноподібних виростів оболонки у внутрішню порожнину клітини. Розростаючись, цистоліти можуть істотно збільшувати розміри клітин, в яких вони розташовані. Основою цистоліта є целюлоза з пектиновими речовинами, які просочені карбонатом кальцію або оксидом кремнію. Утворення цистолітів пов’язане з особливостями обміну речовин і мінерального живлення, як це спостерігається при утворенні кристалічних включень в рослинних клітинах.
Вторинні хімічні зміни клітинної оболонки.
Процес диференціації рослинних клітин супроводжується також різноманітними вторинними змінами їхнього хімічного складу та будови відповідно до тих функцій, які розпочинає виконувати клітина в рослинному організмі. До основних типів вторинних змін клітинних оболонок належать хімічні модифікації щляхом просочування – інкрустації або нашарування – адкрустації певними хімічними речовинами. Залежно від хімічної природи речовин, клітинні оболонки можуть піддаватися:
– здерев’янінню – лігніфікації;
– окорковінню;
– кутикізації;
– ослизненню;
– мінералізації.
Здерев’яніння – лігніфікація.
При здерев’янінні оболонка залишається прозорою і безбарвною, але ущільнюється, стає твердішою, менш еластичною і крихкою. Процес здерев’яніння оболонки зумовлюється утворенням у ній особливої речовини – лігніну. Хімічна природа лігніну досить складна, він належить до полімерних продуктів ферментативної конденсації фенольних сполук. Як і целюлоза, лігнін не містить азоту і має менше атомів кисню. На відміну від целюлози, лігніну не властиве подвійне променезаломлювання.
Мікроскопічні, поляризаційні та рентгенографічні дослідження свідчать про те, що лігнін є лише матриксом, що заповнює міжфібрилярні проміжки целюлозних оболонок. Особливо характерним є роздерев’яніння кам’янистих клітин у достигаючих груш та айви. Це помітили вчені у деревині плодових культур, пошкоджених морозами.
Кількість лігніну у різних деревних порід неоднакова:
– у хвойних – до 30%;
– у листяних – до 25%;
– в оболонці горіхів – до 60%.
Включення лігніну в оболонку клітини спричиняє в ній ряд змін: потовщення, ущільнення, підвищення питомої маси, більший показник заломлення світла, також змінюються фізичні властивості самої оболонки. Вона стає твердішою, більш крихкою і менш еластичною порівняно з целюлозою. Часто спостерігається зв’язок між кількістю лігніну в деревині та її твердістю. Наприклад, м’яка деревина тополі має 21% лігніну, а дуба – 28%. Здерев’янілі оболонки вбирають менше води і при висиханні менше, ніж целюлозна оболонка, зменшуються в об’ємі та деформуються.
Лігнін в оболонках відкладається нерівномірно.
Фрагмент молекули лігніну
Найшвидше дерев’яніє серединна пластинка оболонки, потім вторинні шари, а третинні, внутрішні, майже не дерев’яніють.
Процес здерев’яніння оболонок ксилеми відбувається досить швидко після відокремлення молодих клітин від камбію. Характер структурно-функціонального взаємозв’язку між лігніном і целюлозою подібний по принципу конструкції залізобетонних споруд: целюлоза – це сталеві арматурні елементи залізобетону, а лігнін заповнює проміжки між цим каркасом. Таким чином, утворюється єдине монолітне ціле.
Фізіологічне значення здерев’яніння в тому, що воно надає більшої механічної міцності оболонкам, тканинам та органам рослини. Воно має також антисептичне значення, так як лігнін консервує, ущільнює оболонку, особливо у відмерлих клітин. Порівняно із целюлозою, лігнін більш хімічно інертний і стійкіший до пошкодження бактеріями, грибками та ферментами.
Окорковіння.
При окорковінні оболонка просочується особливою речовиною – суберином – високополімерною речовиною, до складу якої входять різні жирні оксикислоти та їхні солі. Просочуючи оболонку, суберин робить її непроникною для води і газів та стійкою проти гниття та дії різних кислот. Завдяки цим властивостям коркову тканину широко використовують для закупорювання різних рідин, що містять гази (пиво, шампанське вино, мінеральні води). Навіть при великому тиску газів корок не пропускає їх.
Інкрустуючи оболонку, суберин, як гідрофобна речовина, ізолює протопласт від надходження поживних речовин. Окорковіла клітина руйнується, в ній залишаються тільки повітря та зруйнований протопласт у вигляді бурої безформної маси, від якої вся тканина має такий самий колір.
Окорковіння властиве переважно клітинам покривної тканини стебла і кореня. Воно зустрічається також в оболонках клітин шкірки насіння, плодів, також після листопаду на листкових рубцях. При пораненні бульб картоплі корковіють оболонки поверхневих клітин, що перетворюються на такий же захисний шар корка, як і на непошкодженій ділянці бульби.
Суберин відкладається в клітинній оболонці і утворює прошарок у вторинному шарі клітинної стінки. Крім суберину до складу оболонок корка входять інші речовини флавоноїдної природи, які перейшли сюди після відмирання протопласта. Ці сполуки доповнюють захисні властивості коркової тканини, бо характеризуються бактерицидною та фунгіцидною дією, що важливо для запобігання інфекції внутрішніх тканин багаторічних органів, для ефективної роботи сочевичок.
Мінералізація.
Досить часто клітинну оболонку просочують різні мінеральні сполуки: карбонат кальцію у водоростей, кремнезем у злаків та хвощів. Мінералізація характерна майже для всіх рослинних клітин, навіть молода клітинна оболонка містить певну кількість солей. Особливо інтенсивно цей процес відбувається у більш старих клітинах і пов’язаний з відкладанням значної кількості мінеральних речовин зовні або по всій товщі оболонки. З віком кількість мінеральних речовин в оболонці збільшується, особливо у хвощів та харових водоростей.
Якщо пагони хвоща прожарити, залишається крем’яний скелет. Карбонат кальцію відкладається у багатьох вищих рослин на поверхні оболонок, а у багряних водоростей він утворює товстий покрив на зовнішній поверхні рослини.
Кутинізація.
Найбільш поширеною групою речовин, що утворюються в клітині і відкладаються за типом адкрустації, є кутин. Сам процес хімічної модифікації оболонки за його участю називається кутинізацією. Кутин за хімічними властивостями подібний до суберину, але відрізняється від нього переважанням насичених оксикислот жирного ряду. Кутин утворює суцільну поверхневу плівку, яка вкриває зверху клітини епідерми усіх надземних органів рослин і межує з повітрям. Ця плівка називається кутикулою. Кутин може просочувати і частину целюлозних глибинних шарів оболонки.
Досить часто плівка кутину на оболонках поверхневих клітин соковитих плодів – винограду, слив, кавунів або листків деяких рослин може містити і певну кількість воску, який теж секретується із цитоплазми, доповнюючи захисні властивості кутикули.
Кутикула стійка до різних реактивів. Вона не розчиняється навіть у міцній сірчаній кислоті, в також стійка до дії різних мікроорганізмів та грибів. Кутинізовані клітинні стінки практично непроникні для води та інших рідин. Тому кутикулярна плівка утворюється на нижніх клітинах, щоб зберегти їх від висихання. Якщо стінки клітини кутинізуються з усіх боків, то таке явище, як і при окорковінні, спричиняє відмирання клітини.
Кутинізація має велике значення при захисті рослин від надмірного випаровування, пошкоджень і проникнення інфекції. Через кутикулу важко проходять речовини, тому транспірація, живлення і газообмін у рослин майже повністю відбувається крізь продихи. Але крізь кутикулу можуть проходити деякі інсектициди, фізіологічно активні речовини, поживні солі, що наносяться шляхом обприскування.
Ослизнення оболонок.
Явище ослизнення властиве лише деяким типам клітин. Оболонка при цьому дуже набрякає у воді внаслідок перетворення геміцелюлози, пектинових речовин або кальози на слиз або камедь. Слиз та камедь є високомолекулярними вуглеводами, що належать в основному до пентозанів та їхніх похідних.
Ослизнення буває нормальним явищем або патологічним процесом. Постійне ослизнення характерне для деяких водоростей, а також для насіння вищих рослин – льону, айви, хрестоцвітих рослин. Ослизнення оболонки сприяє проростанню насіння, бо таке насіння прилипає до грунту. Крім того, утворений на поверхні насіння слиз затримує воду біля молодого проростка і захищає його від висихання. Здатність до ослизнення насіння льону та гірчиці використовується з лікувальною метою.
Патологічний характер ослизнення – гумоз – зустрічається у багатьох представників кісточкових плодових. Після дощової погоди в цих рослин з кори виділяються набряклі слизові маси, які, застигаючи, утворюють тверді напливи камеді. Це явище особливо помітне також при ушкодженні кісточкових морозами. Інтенсивний гумоз персиків та абрикос майже завжди є показником хворобливого стану цих дерев.
Охарактеризовані зміни, що відбуваються в оболонках рослинних клітин, мають важливе значення. Вони тісно пов’язані з певним напрямком біохімічних процесів, зумовлюють відповідний рівень життєдіяльності рослини, а ряд змін клітинної оболонки має пристосувальне значення. Здерев’яніння, окорковіння, ослизнення, мінералізація та кутинізація зумовлюють захисні функції клітинної оболонки, надають їй міцності, пружності, хімічної стійкості та інших властивостей, що у своєму комплексі забезпечують оптимальну життєдіяльність рослинного організму. Зміни властивостей клітинної оболонки є пристосувальними реакціями, що виникли протягом історичного розвитку рослинних організмів і тісно пов’язані з умовами їхнього життя.
Гумоз – хворобливе перетворення оболонок в стовбурі вишні, сливи в звязку з нанесеними ушкодженнями.
|
В оболонці є непотовщені місця – пори (у первинній оболонці вони називаються поровими полями), через які здійснюється зв’язок між сусідніми клітками. Крізь порові поля і пори проходять тонкі тяжи цитоплазми – плазмодесми, які зв’язують цитоплазму сусідніх кліток. По них здійснюється обмін речовинами між сусідніми клітками. Плазмодесми разом з елементами провідної тканини сполучають клітки і тканини організму в єдине ціле.
Обмін речовин дозволяє клітинам впливати на розвиток і роботу одна одної, і кожна тканина впливає на життєдіяльність всіх інших тканин. Цим створюється координація роботи всіх частин єдиного організму цілої рослини.
Оболонка може просочуватися і мінеральними речовинами – переважно солями кальцію і сіліціуму.
|
|
Бувають, проте, в життя клітин і такі моменти, коли їх оболонка повинна стати менш міцною, ніж була до цього. Це відбувається, наприклад, при проростанні насіння.
Отже, полісахаридна клітинна оболонка – характерна риса будови рослинної клітки клітини , що відрізняє її від тваринної клітини.
Оболонка, або клітинна стінка – це захисне утворення. Під оболонкою знаходиться цитоплазма. Зовнішній її шар, що примикає до оболонки, – це мембрана плазмалема. Вона є комбінацією шарів жироподібних і білкових молекул. Такі мембрани називаються ліпопротеїновими («ліпос» жир, «протеїн» білок). Мембрана такої конструкції відмежовує також цитоплазму від вакуолей і називається тонопластом. Багато органоїдів клітини побудованів з ліпопротеинових мембран. Проте в кожному випадку мембрана побу із дована з жироподібнихх речовин (ліпідів) і білків, властивих саме даній мембрані. Якісна різноманітність ліпідів і особливо білків колосальна, звідси величезна різноманітність мембран, що відрізняються за властивостями
|
Плазмалема регулює надходження речовин в клітину, забезпечує їх вибіркове проникнення. Добре проникають через неї вода і гази. При хімічних або фізичних змінах в зовнішньому і внутрішньому середовищі клітинні мембрани змінюють свою проникність, а також ступінь і сам характер її вибірковості. На цьому ґрунтуються механізми регуляції руху речовин в клітину і з клітини.
.У типовій рослинній клітині є крупна вакуоля, виповнена рідким вмістом. Часто вакуоля займає майже весь об’єм клітини, так що цитоплазма складає лише тонкий шар, прилеглий до клітинної оболонки. У молодих клітин буває декілька дрібних вакуолей, які у міру розвитку клітини розростаються і зливаються в одну. Вміст вакуолі – клітинний сік – це водний розчин дуже багатьох речовин: цукрів, амінокислот, інших органічних кислот, пігментів (фарбувальних речовин), вітамінів, дубильних речовин, алкалоїдів, глікозидів, неорганічних солей (нітратів, фосфатів, хлоридів), іноді також білків.
|
ВКЛЮЧЕННЯ
|
![]() Мал. Кристали щавлевокислого кальцію в вакуолях клітин.
Мал. Кристали щавлевокислого кальцію в вакуолях клітин.
Зліва – в клітинах черешка листка бегонії королівської. Справа – в клітці ряски малої. 1 – крохмальні зерна; 2 – друзи; 3 – ядро; 4 – рафіди; 5 – вакуоля; 6 – цитоплазма.
ЗАПАСНІ ВКЛЮЧЕННЯ РОСЛИННОЇ КЛІТИНИ — тимчасові утворення клітини, які виникають чи зникають у процесі її життєдіяльності і належать до ергастичних речовин (рисунок). З.в.р.к. — вуглеводи, білки і жири. Основне місце їх локалізації — цитоплазма та вакуолі. Речовини, не розчинні у воді, утворюють оформлені включення, що мають вигляд зерен, кристалів і крапель, а розчинні — зазвичай рідкі неоформлені включення. З.в.р.к. входять до складу багатьох ЛП, а ознаки їх будови є видоспецифічними і використовуються при аналізі ЛР і сировини в порівняльному анатомічному аналізі.
Внаслідок фотосинтезу в клітинах зелених рослин утворюються органічні речовини, частина яких відкладається про запас. В ролі запасних поживних речовин зустрічаються основні групи органічних сполук – вуглеводи, ліпіди та білки.
Вони нагромаджуються у плодах та насінні, у коренях, стеблах, бульбах та кореневищах. Під час ростових процесів ці речовини включаються в обмін речовин як джерело енергії та метаболітів.
Різні форми запасних поживних речовин належать до категорії включень – тимчасових компонентів клітин, здатних утворюватись і ферментативно розкладатись у різні періоди їхньої життєдіяльності.
Вуглеводи.
До основних запасних вуглеводів належить крохмаль. Це один з найпоширеніших полісахаридів, який відкладається в усіх рослинах, крім грибів та ціанобактерій. За фізіологічним призначенням і місцезнаходженням, крохмаль розрізняють трьох типів: асиміляційний, транзиторний та запасний.
Асиміляційний крохмаль у вигляді дрібних зерняток утворюється у хлоропластах мезофілу листка, молодих стебел та інших хлорофілоносних клітинах, в яких відбувається інтенсивний фотосинтез.  Первинний крохмаль довго не затримується в хлоропластах, швидко гідролізується до розчинних сахаридів, які потім транспортуються до місць споживання.
Первинний крохмаль довго не затримується в хлоропластах, швидко гідролізується до розчинних сахаридів, які потім транспортуються до місць споживання.
Транзиторний крохмаль утворюється тимчасово на шляхах переміщення сахаридів до місць відкладання про запас. Багато крохмалю цих двох типів утворюється в листках картоплі, гороху, бобів, менше у гречкових, гвоздикових, а найменше у лободових. З однодольних рослин мало крохмалю утворюється в листках злаків і майже не виявлено його у лілійних та орхідних.
Запасний крохмаль нагромаджується у великих кількостях в спеціалізованих запасаючих тканинах та органах – ендоспермі, сім’ядолях, паренхімних клітинах деревини стебла та кореня, бульбах, цибулинах, кореневищах, звідки він поступово мобілізується для життєвих потреб рослинного організму. Запасний крохмаль відкладається у різних рослин у вигляді зерен різної форми та розмірів. У багатьох рослин крохмальні зерна мають добре виражену шаруватість внутрішньої структури, зумовлену чергуванням шарів крохмалю різного ступеня оводненості. Точка, навколо якої розміщені шари, називається ядром крохмального зерна. Залежно від розташування ядра, крохмальні зерна поділяють на концентричні (у пшениці та жита) та ексцентричні (у картоплі). Зерна з ексцентричною шаруватістю утворюються і розвиваються біля поверхні лейкопластів, а з концентричною – у центрі строми пластиди.
Крім простих крохмальних зерен, бувають зерна складні і напівскладні. Складні крохмальні зерна зліплено з кількох зерняток – двох, трьох і більше. Напівскладні зерна відрізняються від складних кількома суцільними зовнішніми шарами крохмалю. Розмір крохмальних зерен дуже різноманітний у окремих рослин:
– у картоплі вони діаметром 70-100 мкм;
– у квасолі – 60 мкм;
– у пшениці – 30-45 мкм;
– у кукурудзи – 15-35 мкм;
– у рису – 3-10 мкм.
Найбільшими є крохмальні зерна у петрового хреста – до 300 мкм діаметром. Величина крохмальних зерен має важливе значення для харчової промисловості, фармакології…
Утворившись у листку, асиміляційний крохмаль гідролізується і у вигляді розчинних сахаридів пересувається у різні частини рослин. У стовбурі дерев він відкладається в живих клітинах усіх тканин, найбільше в деревній паренхімі, серцевинних променях та флоемній паренхімі. Багато крохмалю нагромаджується в коренях багаторічних рослин, а також в насінні. Найбільше крохмалю в різних тканинах стебла та кореня виявляється восени.
У кожній клітині крохмальні зерна тісно пов’язані з іншими компонентами клітинних включень. Крохмальних зерен в клітині, де вони відкладаються як запасні речовини, може бути так багато, що вони заповнюють всю клітинну порожнину. В таких клітинах від цитоплазми залишається тільки тоненька сіточка між окремими зернами. У різних рослинах нагромаджується неоднакова кількість крохмалю. Група рослин, у яких відкладається багато крохмалю, називаються крохмаленосними. У клітинах бульб картоплі крохмалю може нагромаджуватись до 25% сирої маси бульб. Багато крохмалю міститься у зернівках злаків, сім’ядолях бобових.
Другим запасним полісахаридом є інулін. Він властивий представникам родини айстрових, зокрема роду Inula, звідки і одержана назва. Інулін міститься в розчиненому стані переважно в клітинному соку паренхімних клітин зимуючих органів та у бульбах топінамбура, жоржини, у коренях кульбаби, цикорію, полину, лопуха… Він замінює в цих рослинах крохмаль. Під мікроскопом інулін у клітинах майже непомітний і являэ собою сферичної форми скупчення голкоподібних кристалів.
У деяких рослин запасними вуглеводами є інші водорозчинні сахариди – сахароза, глюкоза, фруктоза – у цукрових буряків, цукрової тростини, соковитих плодах.
Рисунок. З.в.р.к.: 1 — прості; 2 — складні крохмальні зерна бульби картоплі (Solanum tuberosum); 3 — cкладні алейронові зерна у насінні рицини звичайної (Ricinus communis)
Найбільше значення серед запасних вуглеводів має крохмаль — органічна сполука групи полісахаридів. Первинний або асиміляційний крохмаль утворюється в хлоропластах у процесі фотосинтезу і має вигляд гранул або зерен. При відсутності фотосинтезу він швидко гідролізується до глюкози і транспортується в інші частини клітини, де може утворювати в лейкопластах (амілопластах) зерна вторинного транзиторного, зберігального або запасного крохмалю. Транзиторний крохмаль утворюється тимчасово на шляхах переміщення глюкози до тканин або органів, що формують його запаси, і не відкладається у великих кількостях. Зерна зберігального крохмалю містяться в клітинах кореневого чохлика та ендодерми стебла, зумовлюючи позитивний гео- та геліотропізм відповідно. Запасний крохмаль у вигляді зерен, як правило, у великих кількостях накопичується в насінинах, бульбах та кореневищах. До ЛР, які є джерелом промислового отримання крохмалю, належать злаки (рис, кукурудза, пшениця), а також картопля, батат тощо.
Ріст крохмального зерна відбувається шляхом нашаровування темних, щільних — денних і світлих, обводнених — нічних шарів крохмалю навколо центру утворення. Крохмальні зерна бувають концентричними — центр утворення співпадає з геометричним центром (пшениця, кукурудза); ексцентричними — центр утворення не співпадає з геометричним центром (картопля); простими — мають один центр утворення (пшениця), складними — центрів утворення два або більше, кожен з яких оточений лише індивідуальними шарами крохмалю; напівскладними — центрів утворення два і більше, а крім індивідуальних, є ще й зовнішні спільні шари крохмалю; складно-напівскладними — утворені з 2 частин, з яких одна — просте зерно, друга — напівскладне (останні два типи зустрічаються у картоплі). Видоспецифічними ознаками крохмальних зерен є їх форма (лінзоподібна, овальна, яйцеподібна, веретеноподібна тощо), яка може залежати від положення в просторі; розміри, тип (прості, складні, ексцентричні, концентричні), характеристика шаруватості (добре виражена, погано виражена, невиражена тощо); наявність центру утворення та його характеристика (у вигляді точки, тріщинки тощо), кількість зерняток у складному зерні. Під дією розчинів йоду крохмальні зерна забарвлюються в синьо-фіолетовий колір. Крохмаль широко використовується як присипка, складова багатьох твердих і м’яких лікарських форм, зв’язувальна і опудрювальна речовина у виробництві таблеток, обволікаюча речовина, джерело глюкози тощо. Запасним полісахаридом рослинної клітини є також інулін. У розчиненому стані він міститься у вакуолях клітин підземних органів рослин родини айстрових, дзвоникових, деяких видів цибулевих, гіацинтів. Під дією α-нафтолу клітини з інуліном забарвлюються у фіолетовий колір; 96° спирт сприяє утворенню сферокристалів інуліну, які бувають різного розміру і здатні набухати. Інулін використовується як джерело D-фруктози, замінник крохмалю та цукру при діабеті, для нормалізації вуглеводного і ліпідного обміну, імунного статусу, як ентеросорбент, має біфідогенну активність. Багатими на інулін є бульби топінамбура, корені кульбаби, цикорію, оману тощо. У деяких рослин запасними вуглеводами є водорозчинні сахариди — глюкоза, фруктоза, сахароза (ягоди винограду, кавуна, коренеплоди цукрового буряка тощо).
Білкові З.в.р.к., як правило, накопичуються у вигляді алейронових або протеїнових зерен. Це тверді безбарвні округлі або еліптичної форми тільця величиною від 0,1 до 20 мкм, які накопичуються переважно в насінні. Напр. у бобових вони знаходяться в клітинах сім’ядоль серед крохмальних зерен; у рицини складають основну масу ендосперму; в зернівках злаків розміщені під плодовою оболонкою, утворюючи зовнішній алейроновий шар клітин ендосперму. Алейронові зерна утворюються при висиханні вакуолей клітин, багатих на ергастичні білки, але при проростанні насінини відбувається зворотний процес — набухання і перетворення алейронових зерен на вакуолі. Алейронові зерна складаються з мембранної (білкової) оболонки й аморфної білкової маси, в якій може бути три типи включень: глобоїди, кристалоїди та кристали оксалату кальцію. Глобоїд — блискуче безбарвне аморфне тільце округлої, іноді гроновидної форми, що складається в основному з фітину — кальцієвої і магнієвої солі інозитфосфорної кислоти. Кристалоїд — білок, який має форму кристала, але у воді набухає. Розрізняють такі типи алейронових зерен: з глобоїдами (насіння бобових, гірчиці, соняшника, зернівки злаків); з глобоїдами і кристалоїдами (насіння льону і рицини); значно рідше зустрічаються зерна з кристалами оксалату кальцію (насіння селерових і винограду). У деяких видів (картоплі, квасолі) утворюються окремі кристалоїди, не оточені аморфним білком. Алейронові зерна поділяють на прості і складні. Прості мають однорідну структуру, а складні серед аморфного білка містять кристалоїди (один або кілька) та глобоїди (один або кілька).
Наявність алейронових зерен встановлюють за допомогою якісних реакцій, характерних для речовин білкової природи, напр., під дією йоду вони забарвлюються в жовтий колір. Будова алейронових зерен також є видоспецифічною ознакою, за якою можна визначити рослину або рослинну сировину. На основі глобоїдів, які отримують з вичавок коноплі та ін., виготовляють фосфоровмісний препарат «Фітин». З.в.р.к. багатьох рослин представлені жирами, що локалізуються, як правило, в насінні і накопичуються у вигляді безбарвних або жовтуватих ліпідних крапель і гранул.
Жири утворюються в цитоплазмі або лейкопластах (олеопластах) і накопичуються в твердому або частіше рідкому стані (олії). У рослин помірних і північних широт утворюються переважно рідкі олії, у тропічних рослин — тверді (насінини кокосової пальми, какао, мускатного горіха).
Рослинні олії входять до складу основ мазей і супозиторіїв, до препарату Лінетол, використовуються для профілактики атеросклерозу, як проносна речовина. У фармацевтичній практиці використовують рідкі олії (плодів оливкового дерева, насіння мигдалю, арахісу, рицини тощо) і тверді (плодів кокосової та олійної пальм, шоколадного дерева (какао)) тощо.
Лінетол
Всі ці речовини – продукти життєдіяльності клітини. Одні з них зберігаються у вакуолярному (клітинному) соці як запасні речовини і з часом знов поступають в цитоплазму для використання. Інші є кінцевими продуктами обміну речовин, що виводяться з цитоплазми. Так, у вакуолю виводиться щавлева кислота; у вакуолярному соці часто відкладаються кристали щавлевокислого кальцію: іноді у формі поодиноких кристалів, в інших випадках у вигляді конгломерату кристалів цієї солі: багатогранних (друзи) або голчастих (рафіди).
|
Різні форми кристалів карбонату, оксалату кальцію і інших солей є важливою діагностичною ознакою анатомічної будови різних систематичних груп рослин.
Друзи оксалату кальцію – характерна ознака кореня ревеню при його мікроскопічному аналізі.
Ефірні олії відносять до кінцевих продуктів метаболізму. Вони можуть накопичуватися в зовнішніх і внутрішніх видільних структурах рослинниого організму.
Приклади якісних мікрореакцій на основні види запасних включень рослинних клітин.
|
Досліджувана речовина
|
Реагент |
Результат (зміна забарвлення) |
|
крохмаль |
розчин Люголя |
синьо-фіолетове |
|
білок |
HNO3 конц. |
жовте |
|
жирна олія |
судан ІІІ |
оранжеве |
|
ефірна олія |
судан ІІІ |
оранжеве |
|
целюлоза |
Cl-Zn-J |
синьо-фіолетове |
|
кутин |
судан ІІІ |
оранжеве |
|
суберин |
судан ІІІ |
оранжеве |
|
лігнін |
HCl конц. |
малинове |
|
антраценпохідні |
NaOH |
рожево-малинове |
|
дубильні речовини |
залізо-амонійні галуни |
темно-зелене (або темно-синє) |
У клітинах поряд із вуглеводами містяться запасні поживні речовини білкового типу. Запасні білки порівняно з конституційними білками, які входять до складу цитоплазми та її структурних компонентів, мають меншу молекулярну масу та інший амінокислотний склад. Найчастіше вони зустрічаються в клітинах у вигляді аморфних мас, дрібних зерняток або кристалоподібних утворень. Тому їх називають кристалоїдами.
Білкові кристали містяться у клітинах багатьох рослин і мають форму правильних кристалічних утворень. У клітинах картоплі кристалоїди лежать у поверхневих шарах, де мають форму правильного кубика. Білкові кристали локалізуються безпосередньо в цитоплазмі, у клітинному соку, а інколи в ядрі
Органічні кислоти. Зумовлюють кислу реакцію клітинного соку. Найчастіше зустрічаються щавлева кислота, яка з’єднується з кальцієм і утворює нерозчинну у воді сіль оксалату кальцію. Вона утворюється як один із побічних продуктів дихання.
Яблучна кислота є найбільш розповсюдженою в різних плодах (яблука, горобина, вишня. Помідори, брусниця, журавлина) зумовлюючи їх кислуватий смак.
Лимонна кислота характерна для плодів лимона (6%), лимонника (10%), журавлини (3%).
Вишня-кислота переважає в плодах винограду (0,3%) і малини.
Бензойна кислота зустрічається в плодах брусниці (800,1%), журавлини. Для неї характерні консервуючі властивості, що зумовлює здатність цих плодів зберігатись як у натуральному так і в замоченому вигляді.
Органічні кислоти підтримують тугор клітини.
Глікозиди – складні органічні речовини, до складу яких входить глюкоза або ін. Цукор, зв’язаний по типу складних ефірів з несахаристими речовинами (аглюконами). Глікозиди, особливо серцеві – отруйні сполуки, які зумовлюють отруйність всієї рослини.
У медицині глікозити мають досить широке застосування. Наприклад, глікозиди таких рослин, як конвалія, наперстянка, горицвіт, застосовують при серцевих захворюваннях. Глікозид амигдалін під впливом ферменту емульсину розщеплюється на глюкозу, безолійний альдегід і дуже отруйну синильну кислоту. Він знаходиться в насінні гіркого мигдалю, тому не насіння не можна використовувати як їжу, бо кілька насінин можуть спричинити сильне отруєння. Амигдалін є також у насіннях сливи, вишні, абрикоса, персика, яблуні, груші, але в невеликих кількостях, то обумовлює їх гіркуватий смак.
Гіркоти – речовини глікозидної природи, вони характерні для полину, золототисячника, тирличу. Гіркоти використовують для поліпшення апетиту і покращення травлення.
Таніди (або дубильні реч.). Особливістю їх є в’яжучий смак, кисла р-а, здатність осаджувати більше і алкалоїди. Таніди містяться в значних кількостях в усіх органах і частинах рослин. Наприклад, у дубових голок їх вміст досягає 75%, у корі дуба – 20%, верби – 13%. Вміст дубильних речовин у плодах під час їх зберігання, достигання, а також внаслідок дії низьких tо знижується. У медицині мають широке застосування. І використовують як в’яжучий засіб при запаленні слизових оболонок, шлункових маткових кровотеч ін.
Пігменти – безбарвні розчинні речовини. Найпоширенішими рослинними пігментами є антоціани і флавони. Вони знаходяться в пелюстках квіток, у листках, стеблах, плодах і насінні. Антоціани органічні сполуки, здебільшого феномені глікозиди. Їх забарвлення залежить від реакції клітинного соку. Якщо останній має кислу реакцію, актоціани набувають червоного кольору різних відтінків, при нейтральній реакції – фіолетового, а при лужній – синього або блакитного. У рослинах реакція клітинного соку може змінюватися, а тому і забарвлення рослин також змінюється.
Наприклад: у медунки на початку цвітіння пелюстки квіточок червоні, з часом їх забарвлення змінюється на фіолетове, а в кінці цвітіння вони синіють. Флавон – пігмент жовтого чи оранжевого кольору. Звичайно на нього багато пелюстки жовтих жоржин, маку, жовті і оранжеві плоди мандаринів, лимонів, апельсинів.
Пігменти відіграють велику біологічні роль. Забарвлюючи квітки, вони приваблюють комах і таким чином сприяють перехресному запиленню. Антоціани вбирають сонячне промінні і сприяють підвищенню tо клітин, що важливо в холодні періоди, при весняних похолодання. Антоціанам притаманні фітонцидні властивості.
Вітаміни – солодкі органічні сполуки різного складу. Вважають, що вітаміни відіграють роль каталізаторів в організмі і беруть участь у синтезі деяких ферментів у клітині. Vit., які розчиняються у воді (В, С. Р) знаходяться в клітинному соці. Решта (розчинних в олії) міститься в цитоплазмі.
Vit. гр. В є:
· В1 (тіамін),
· В2 (рибофлавін),
· РР (нікотинова кислота, фолієва кислота, (Вс)
· і В12 (ціанокоболомін).
Vit. В1 – міститься в пивних і мінеральних дріжджах, зародках і зовнішніх оболонках насіння злаків: бобових, у гречаних і вівсяних крупах шипшині, білокачанній капусті, моркві, картоплі, зеленому горошку. Нестача Vit. В1 в організмі спричинює нервове захворювання бері-бері.
Vit. В2 – міститься в дріжджах, горосі, цибулі, в зародках пшениці, кукурудзі і ін. Його нестача в організмі призводить до дерматитів запалення язика і губ, послаблення зору.
Vit. РР (В5) У значних кількостях міститься в дріжджах, грибах, помідорах, картоплі, моркві, гречаній крупі, арахісі, кропиві, квасолі, шпинаті і ін. Його нестача приводить до захворювання на пелагру, що характеризується нервовим інтенсивним розладом, запаленням слизової оболонки рота і язика, полосами, появою плям на шкірі.
Фолієва кислота (Вс) міститься в дріжджах, листях шпинату, салату, зеленої цибулі, щавлю, петрушки, селери, в суницях. Нестача в організмі є причиною недокрів’я (анемії).
Vit. В12 В основному міститься в продуктах тваринного походження – печінці і ін. внутрішніх органах. Нестача спричинює порушення координації руху, шлунково-кишкових розладів, запальні процеси в ротовій порожнині, втрату апетиту.
Vit. С (аскорбінова кислота) дуже поширений у рослинах. Найбільше його міститься у плодах шипшини, чорної смородини, червоного перцю, в корінні хрону, листках петрушки, щавлю, шпинату, салату, кропиви, капусти всіх видів, кабачках, порічках, кизилі, обліписі, помідорах, суницях. Vit. С підвищує імунітет, його нестача спричиняє захворювання цингою.
У клітинному соці містяться також токсини – дуже отруйні сполуки, що утворюються в процесі життєдіяльності рослин. Наприклад, токсин рицин знаходиться в насінні рицини, токсин робін – у корі білої акації. Тому, не знаючи рослини, не можна її не лише їсти, а й жувати, бо це може призвести до отруєння і навіть смерті.
Фітонциди – рідкі або леткі сполуки, отруйні для бактерій, найпростіших, грибів, деяких видів вищих рослин, комах, молюсків, земноводних і не отруйні для людини.
Фітонциди притаманні лише насінним рослинам і проявляють вибіркову дію. Легкі Фітонциди мають велике значення при озелененні міст. Повітря яких наповнене не хвороботворними бактеріями. Фітонциди застосовують для лікування гнійних запальних процесів шкіри, карбункулів, стоматитів, опіків, використовують проти стафілококів, патогенних грибів, кишкової палички, дизентерії, бруцельозу.
Ефі́рна олі́я (етерні олії, етери) — рідка летка суміш органічних речовин, котрі виробляються рослинами і надають їм запаху.
Більшість ефірних олій добре розчиняється в бензині, ефірі, хлороформі, ліпідах, в оліях та інших ліпофільних речовинах, та погано — у воді.
Ефірні олії в залежності від способу добування поділяються на натуральні, натуралізовані і синтетичні. Лікувальні властивості має лише натуральна ефірна олія. Синтетична ефірна олія — це продукт фракції нафти або кам’яновугільної смоли.
Більшість відомих людству лікарських рослин є ефіроносами.
Антибіотики – складні органічні сполуки, які утворюють в процесі життєдіяльності клітин нижчих організмів – бактерій (граміцидин), грибів (пеніцилін, аспергілін). Вони згубно діють на організми інших нижчих рослин, що дозволяє використовувати їх у медичній практиці.
Мінеральні речовини – потрапляють у клітину разом з водою. Це різні неорганічні сполуки. Деякі із елементів, що потрапляють у клітину, життєво необхідні рослині (Са, К, N, Р, Fе тощо) ін. не використовуються рослиною і залишаються в клітині.
Значення вакуоль важливе: вони формують внутрішнє водне середовище клітин, з їх допомогою відбувається регуляція водно-сольового обміну, підтримується тургорно-гідростатичний тиск внутрішньо-клітинної рідині, в них накопичуються запасні речовини, відкладаються кінцеві продукти обміну речовин.
Алкалоїди
Кристалічні включення клітин.
У багатьох рослин є тверді включення, серед яких найпоширенішими є нерозчинні кристалізовані солі кальцію, особливо оксалат кальцію. Ці солі утворюються внаслідок обміну речовин і можуть відкладатися у великій кількості. Оксалат кальцію в живих клітинах може відкладатися про запас, який потім, розчиняючись, використовується рослиною в процесі обміну, як резервний фонд іонів кальцію. Наприклад, у цитрусових, складноцвітих. У багаторічних рослин основна кількість оксалату кальцію нагромаджується у старих листках та корі. Утворюються оксалати в основному у вакуолях у вигляді кристалів різної форми. У залежності від форми та величини кристалів вони називаються друзи, рафіди та стилоїди.
У покривних тканинах деяких рослин зустрічаються цистоліти – аморфні гроноподібні утворення карбонової солі кальцію. Тіло цистоліта формується вп’ячуванням оболонки у порожнину клітини, тому клітина, в якій утворюється цистоліт, за своїми розмірами більша, ніж інші. Крім оксалату кальцію у клітинах утворюються кристали виннокислого кальцію, сульфату кальцію, діоксиду кремнію.
Список рекомендованої літератури
а) Основна:
1. Сербін А.Г. Фармацевтична ботаніка: підручник / А.Г. Сербін, Л.М. Сіра, Т.О. Слободянюк / Під ред. Л.М. Сірої. – Вінниця: Нова книга, 2007. – С. 21–28.
2. Пішак В.П. Медична ботаніка. Анатомія рослин з практикумом: Навч.-метод. посібник / В.П. Пішак, В.В. Степанчук / За ред. чл.-кор. АПН України, проф. В.П. Пішака. – Чернівці: Медуніверситет, 2007. – С. 38–60.
3. Нечитайло В.А. Ботаніка. Вищі рослини. / В.А.Нечитайло, Л.Ф. Кучерява. – Київ: Фітосоціоцентр, 2000. – С. 17-20.
4. Ткаченко Н.М., Сербін А.Г. Ботаніка: Підручник. – Х.: Основа, 1997. – 432 с.
5. Григора І.М., Алейніков І.М., Лушпа В.І. та ін. Курс загальної ботаніки. – Київ: Фітосоціоцентр, 2003. – 500 с.
6. Стеблянко М.І., Гончарова К.Д., Закорко Н.Г. Ботаніка: Анатомія і морфологія рослин: Навч. посібник / За ред. М.І. Стеблянка. – К.: Вища школа, 1995. – 384 с.
б) Додаткова:
7. Сербин А.Г., Серая Л.М., Ткаченко Н.М., Слободянюк Т.А. Медицинская ботаника: Учебное пособие для иностранных студентов. – Харьков: Изд-во НФАУ, 2000. – 283 с.
8. Географія рослин з основами ботаніки: Навч. посібник / Гришко-Богменко Б.К., Морозюк С.С., Мороз І.В., Оляніцька Л.Г. – К.: Вища школа, 1991. – 255 с.
9. Жизнь растений / Гл. ред. акад. АН СССР А.Л. Тахтаджян. – М.: Просвещение, 1970-1982. – Т.1–6.
10. Лікарські рослини: Енциклопедичний довідник / Відп. ред. А.М. Гродзинський. – К.: Українська енциклопедія ім. М.П.Бажана, 1992. – 544 с.
11. Рейвн П., Эверт Р., Айкхорн С. Современная ботаника: В 2-х т. – М.: Мир, 1990.
12. Randy Moore, W.D. Clark, Kingsley R.Stern, Darell Vodopich. Botany.– Toronto: WCB, 1995. – 824 р.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}